
Response of leaf functional traits and anatomical structure to altitude in Crataegus songarica K. Koch in Tianshan wild fruit forest
-
摘要:
为揭示准噶尔山楂(Crataegus songarica K. Koch)对海拔高度变化的响应,以新疆伊犁果子沟不同海拔(1100~1700 m)的准噶尔山楂作为研究对象,分析其叶片功能性状及解剖结构差异。结果显示:(1)随着海拔的升高,准噶尔山楂的叶长、叶宽、叶面积、比叶面积、叶绿素含量呈降低趋势,叶长宽比、比叶重、叶干物质含量呈上升趋势,比叶重可塑性最强。(2)随着海拔升高,叶厚、栅栏组织厚度、上下表皮厚度呈上升趋势;主脉厚度、突起度呈下降趋势,主脉突起度可塑性最强。(3)叶面积与叶长、叶长宽比、比叶面积呈正相关;叶厚与上下表皮厚度、主脉突起度等呈正相关。研究结果表明,准噶尔山楂在高海拔区域主要通过增加叶厚和叶干物质含量以提高抗逆性,减少叶面积、叶绿素含量来降低高光强对叶片的伤害;在低海拔区域主要通过增加叶面积、叶绿素含量来促进有机物的积累。
Abstract:This study investigated the adaptive responses of Crataegus songarica K. Koch to varying altitudes (1 100-1 700 m) in Guozigou, Yili, Xinjiang, China, focusing on leaf functional traits and anatomical structure. Results showed that: (1) An increase in altitude led to a reduction in leaf length, leaf width, leaf area, specific leaf area, and chlorophyll content, but an increase in the leaf length to width ratio, specific leaf weight, and leaf dry matter content, with specific leaf weight showing the strongest plasticity. (2) Higher altitudes were associated with increasing leaf thickness, palisade tissue thickness, and upper and lower epidermis thickness, but decreasing central vein thickness and protrusion, with central vein protrusion showing the strongest plasticity. (3) Leaf area was positively correlated with leaf length, length-width ratio, and specific leaf area, while leaf thickness was positively correlated with upper and lower epidermis thickness and central vein protrusion. Overall, our findings indicated that Crataegus songarica adapts to high-altitude conditions by enhancing leaf thickness and dry matter content for better stress tolerance and by diminishing leaf area and chlorophyll content to mitigate leaf damage from intense sunlight. Conversely, at lower altitudes, the plant increases leaf area and chlorophyll content to facilitate organic matter accumulation.
-
叶片作为植物与大气环境接触面积最大的器官,不仅是光合作用的主要场所,更是进行蒸腾作用、呼吸作用等生理活动的基础[1]。因此,植物本身对环境的适应性主要反映在叶片上,其中叶片功能性状和解剖结构最能体现植物对环境的适应性[2]。海拔梯度的变化通常会导致植物物质代谢和功能性状等发生变化[3]。随着海拔的升高,平均气温逐渐降低,降水减少,大气压变低,太阳辐射增强,植物生长季变短,从而导致植物叶片功能性状和解剖结构产生一系列变化[4]。仲小茹等[5]研究表明,米槠(Castanopsis carlesii(Hemsl.)Hayata.)叶片叶面积、长宽比等在高、低海拔之间呈显著差异。Chai等[6]发现秦岭栎树(Quercus acutissima L.)叶干物质含量随海拔呈先上升后下降的趋势。峨眉冷杉(Abies fabri (Mast.) Craib)叶片栅栏组织厚度、海绵组织厚度随海拔升高不断增大[7],黄柏(Phellodendron amurense Rupr.)则通过增加叶片栅栏组织、海绵组织、主脉等的厚度以适应高海拔[8]。薄毛海绵杜鹃(Rhododendron aganniphum var. schizopeplum)主要通过增加叶片角质层厚度、表皮厚度和叶肉厚度来增强对外界极端环境的适应能力[9]。以上研究表明植物在不同环境中有不同的适应机制。因此,了解不同海拔植物的叶功能性状和解剖结构的差异有利于理解物种的分布特征及其适应环境的方式。
目前关于叶功能性状和解剖结构对海拔梯度的响应已有较多研究,但地区不同、物种不同,其变化规律也不尽相同。对不同海拔准噶尔山楂(Crataegus songarica K. Koch)叶功能性状和解剖结构进行分析,将有助于揭示天山野果林生态系统中植物采取的适应方式以及性状进化的环境应力。
天山野果林是世界重要的野生果树天然基因库,蕴含丰富的战略性生物资源[10]。准噶尔山楂作为天山野果林的主要树种之一,对区域生物多样性和生态系统稳定性的维持至关重要。准噶尔山楂为蔷薇科山楂属植物,主要分布于新疆伊犁局部山区[11],其果实可入药[12]。此外,在改善当地生态环境、维持天山野果林的原生性方面具有很好的利用前景[13]。长期以来,由于环境变化和人为活动的干扰,准噶尔山楂栖息地遭到严重破坏,致使其天然资源面积萎缩、病虫害严重、群落退化,严重威胁着该地区生态系统的稳定[14]。叶功能性状和解剖结构是植物为适应环境表现出的功能特征,研究二者相互关系及其随海拔梯度的变化规律有助于理解植物应对环境变化的适应策略[15]。因此,本研究拟通过对天山野果林不同海拔的准噶尔山楂叶片功能性状及其解剖结构的分析,探讨其对海拔变化的响应,为准噶尔山楂资源的保护提供基础资料。
1. 材料与方法
1.1 研究区概况
本研究区域位于新疆伊犁果子沟(44°22′~44°23′N,80°57′~80°58′E),土壤类型为黑钙土,土层较厚,土壤肥力较高[16],年平均气温10.4 ℃,年降水量为260~800 mm,平均日照时数达2898.4 h。果子沟地处伊犁河谷,具有温暖湿润的“海洋性”气侯特色[17]。准噶尔山楂的伴生乔木主要包括新疆野杏(Prunus armeniaca L.)、新疆野苹果(Malus sieversii (Ledeb.) Roem.)等;林下灌木主要有野蔷薇(Rosa multiflora Thunb.)、黑果小檗(Berberis atrocarpa Schneid.)、驼绒藜(Ceratoides latens (J. F. Gmel.) Reveal et Holmgren)等;草本植物主要有卷耳(Cerastium viscosum L.)、牻牛儿苗(Erodium stephanianum Willd.)、扁蓄(Polygonum aviculare L.)、早熟禾(Poa annua L.)、天蓝苜蓿(Medicago lupulina L.)、荨麻(Urtica fissa E. Pritz.)、短距凤仙花(Impatiens brachycentra Kar. et Kir.)等[18]。
1.2 实验方法
1.2.1 材料采集
2022年7月在新疆伊犁果子沟准噶尔山楂分布的海拔范围(1 100~1 700 m)内,海拔每升高100 m设置3个20 m×20 m的样地,共18个样地,作为采样点。每个采样点选择6株长势良好、发育程度相近、健康的成年植株(胸径大于2 cm)[19],从不同方位随机采集10片无蛀虫、无破损的叶片,用刀片截取叶片中部0.5~1.0 cm宽的小段,放入装有FAA固定液(90 mL 70%乙醇 + 5 mL 40%甲醛 + 5 mL冰醋酸)的离心管内,做好标记,密封后放入4 ℃冰箱,用于测量叶片解剖结构。通过GPS定位,记录各采样点的海拔高度、经纬度等信息(表1)。
表 1 样地基本信息Table 1. Basic information of sample plots样地号
Sampling site海拔
Altitude / m纬度
Latitude / N经度
Longitude / E坡度
Slope / °1 1 156 44°22′20.19″ 80°57′43.15″ 13.89 2 1 180 44°22′20.08″ 80°57′40.26″ 16.13 3 1 195 44°22′20.23″ 80°57′41.51″ 17.98 4 1 209 44°22′20.22″ 80°57′44.35″ 18.85 5 1 236 44°22′20.08″ 80°57′44.25″ 19.56 6 1 274 44°22′20.94″ 80°57′45.13″ 20.23 7 1 312 44°22′20.57″ 80°57′49.05″ 20.63 8 1 358 44°22′21.26″ 80°57′50.12″ 21.05 9 1 376 44°22′21.27″ 80°57′51.26″ 22.47 10 1 410 44°22′22.28″ 80°57′52.41″ 23.50 11 1 430 44°22′26.43″ 80°58′15.16″ 24.65 12 1 483 44°22′23.30″ 80°57′54.02″ 24.78 13 1 510 44°22′22.31″ 80°57′55.33″ 25.01 14 1 538 44°22′23.32″ 80°57′56.21″ 25.58 15 1 580 44°22′21.29″ 80°57′59.58″ 27.83 16 1 612 44°22′22.80″ 80°57′43.92″ 29.92 17 1 623 44°22′23.59″ 80°57′45.83″ 30.97 18 1 630 44°23′13.15″ 80°57′37.41″ 31.65 1.2.2 功能性状测定
采用便携式叶绿素测定仪(SPAD-502,Minolta Co,日本)测定叶绿素含量值(Soil and plant analyzer development,SPAD);用EsponV19扫描仪扫描平展叶片的图像,结合ImageJ软件分析测定叶长(Leaf length,LL)、叶宽(Leaf width,LW)、叶面积(Leaf area,LA);利用精度为0.001 g的电子天平称量叶片鲜重(Fresh weight,FW),将样品置于80 ℃恒温干燥箱中烘干至恒重,并称取叶片干重(Dry weight,DW)。通过公式分别计算叶干物质含量(Leaf dry matter content,LDMC)、比叶面积(Specific leaf area,SLA)和比叶重(Specific leaf weight,SLW)。
LDMC=DW/FW (1) SLA=LA/DW (2) SLW=DW/LA (3) 1.2.3 叶片解剖结构
使用常规石蜡制片技术,将固定好的叶片制成永久装片。使用尼康显微成像系统(Nikon MAD 42055)及分析软件(Image Measuring Software Version 5.0)测量解剖结构,包括叶片厚度(Leaf thickness,LT)、上表皮厚度(Upper tissue thickness,UT)、角质层厚度(Cuticle thickness,CT)、栅栏组织厚度(Palisade tissue thickness,PT)、主脉厚度(Main vein thickness,MVT)、下表皮厚度(Lower epidermis thickness,LET)和海绵组织厚度(Spongy tissue thickness,ST)。根据以上指标测量结果,计算栅海比(PT/ST)、叶片组织结构紧密度(Cell tense ratio,CTR)、叶片组织结构疏松度(Spongy ratio,SR)、主脉突起度(Main vein projection,MVP)、可塑性指数(Plasticity index,PI),计算公式如下:
CTR=PT/LT (4) SR=ST/LT (5) MVP=MVT/LT (6) PI = 1−各种群中最小平均值/各种群中最大平均值[20] (7)
1.3 数据分析
通过Excel 2021软件对数据进行处理,使用SPSS 26.0软件对实验数据进行正态检验(S-W检验),然后进行单因素方差分析(One-way ANOVA)和差异显著性检验(Duncan法),图表中数据为平均值±标准误,采用Origin 2021软件制图,采用R语言ggcorr包进行相关性分析。
2. 结果与分析
2.1 不同海拔叶功能性状的变化特征
研究结果发现,随着海拔的升高,叶长、叶宽、叶面积、比叶面积、叶绿素含量均呈下降趋势;叶长宽比、比叶重、叶干物质质量则呈上升趋势。叶长在海拔1 100~1 200 m和1 600~1 700 m处差异显著。叶宽在高海拔段(1 500~1 700 m)与低海拔段(1 100~1 300 m)差异显著。叶长宽比在低海拔段(1 100~1 300 m)差异显著(P<0.05)。叶面积在1 100~1 200 m处与1 300~1 700 m差异显著。比叶面积在1 100~1 200 m处与其他海拔段差异均显著。比叶重在1 500~1 600 m、1 600~1 700 m处与其他海拔差异均显著。叶干物质含量在1 100~1 200 m与1 200~1 300 m差异不显著,但与其他海拔段差异显著。叶绿素含量在1 600~1 700 m与其他海拔段差异显著(表2)。
表 2 不同海拔叶片功能性状变化特征Table 2. Changes of leaf functional traits at different altitudes海拔
A / m叶长
LL / cm叶宽
LW / cm叶长宽比
LL/LW叶面积
LA / cm2比叶面积
SLA / cm2/g比叶重
SLW / mg/cm2叶干物质含量
LDMC / g/g叶绿素含量
SPAD1 100~1 200 5.92±0.15a 5.73±0.16a 1.00±0.04c 17.56±0.97a 170.58±10.22a 6.03±0.47c 0.37±0.03c 42.01±1.51a 1 200~1 300 5.77±0.16ab 5.57±0.12a 1.02±0.04bc 16.19±0.81ab 140.34±8.75b 6.08±0.43c 0.46±0.03bc 40.34±0.87a 1 300~1 400 5.68±0.13ab 5.38±0.17ab 1.10±0.05abc 14.56±0.84bc 130.82±7.88bc 6.75±0.38c 0.49±0.04ab 39.73±0.81a 1 400~1 500 5.59±0.18ab 5.38±0.21ab 1.14±0.04ab 13.77±0.61c 118.24±10.37bcd 7.57±1.23c 0.49±0.03ab 39.28±1.33a 1 500~1 600 5.42±0.19ab 4.98±0.17bc 1.16±0.05a 13.25±0.90c 113.81±7.68cd 11.12±1.08b 0.50±0.05ab 38.93±1.15a 1 600~1 700 5.39±0.18b 4.61±0.12c 1.18±0.04a 7.88±0.59d 96.10±6.81d 14.17±0.69a 0.58±0.03a 34.89±0.89b 注:数据显示为平均值±标准误,同列不同字母表示不同海拔各参数差异显著(P<0.05)。 Notes: Data is shown as Mean±Standard error, and different letters in the same column indicate significant differences in parameters at different altitudes (P<0.05). 2.2 叶片功能性状可塑性和相关性分析
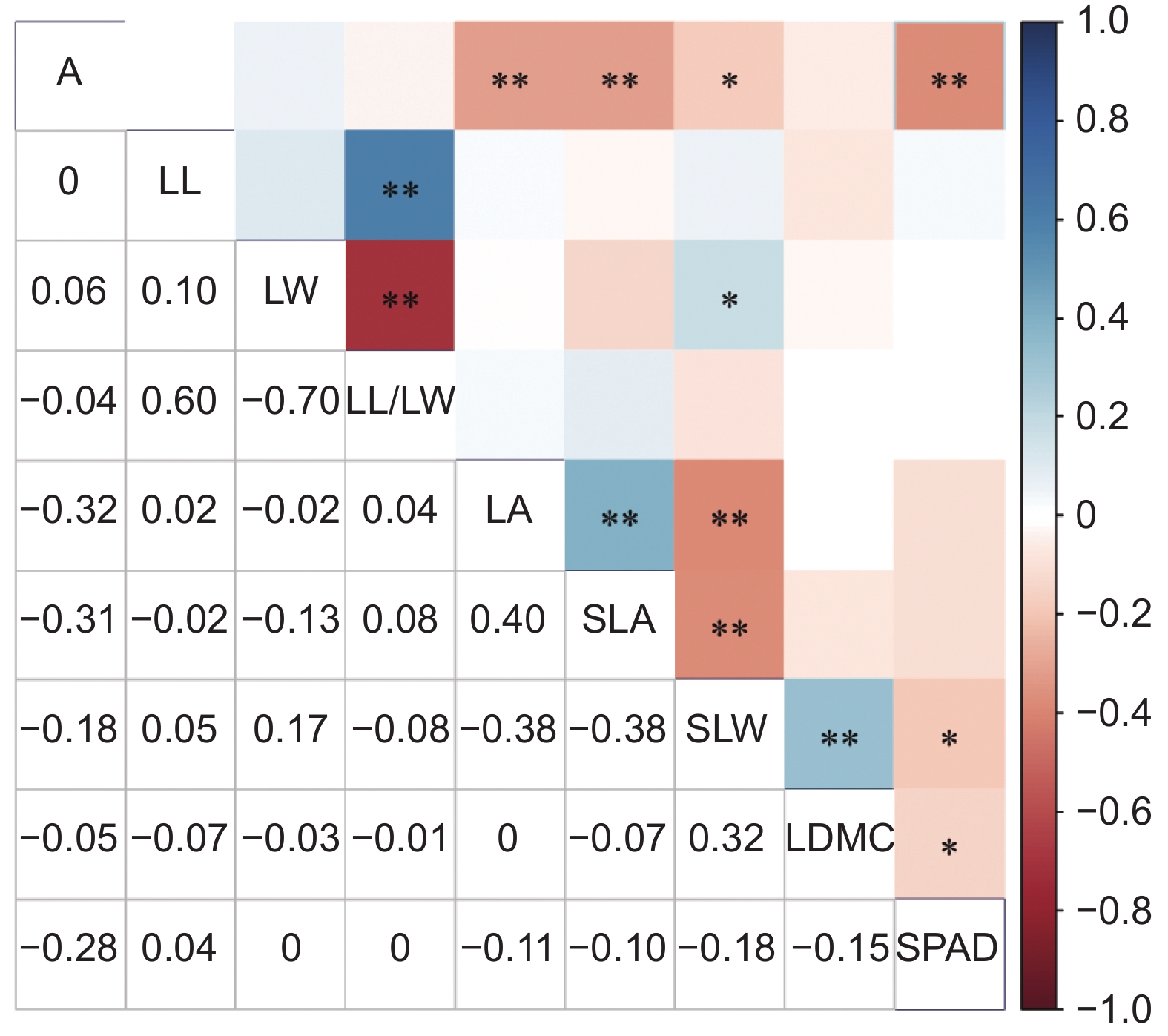
准噶尔山楂叶片功能性状之间的相关性分析结果显示,叶长与叶长宽比呈极显著正相关;叶宽与叶长宽比呈极显著负相关;叶面积与比叶面积呈极显著正相关,与比叶重呈极显著负相关;比叶重与叶干物质含量呈极显著正相关,与叶绿素含量呈显著负相关;叶干物质含量与叶绿素含量呈显著负相关(图1)。
![]() 图 1 准噶尔山楂叶片功能性状间相关性分析Figure 1. Correlation analysis of leaf functional traits in Crataegus songarica*:P<0.05;**:P<0.01. Same below.
图 1 准噶尔山楂叶片功能性状间相关性分析Figure 1. Correlation analysis of leaf functional traits in Crataegus songarica*:P<0.05;**:P<0.01. Same below.通过对准噶尔山楂叶功能性状进行可塑性分析,发现各指标数值范围在0.20~0.60,其中比叶重可塑性指数最大,为0.60,其次是叶干物质含量,为0.51,最小的为叶绿素含量,为0.20。
2.3 不同海拔叶解剖结构的变化特征
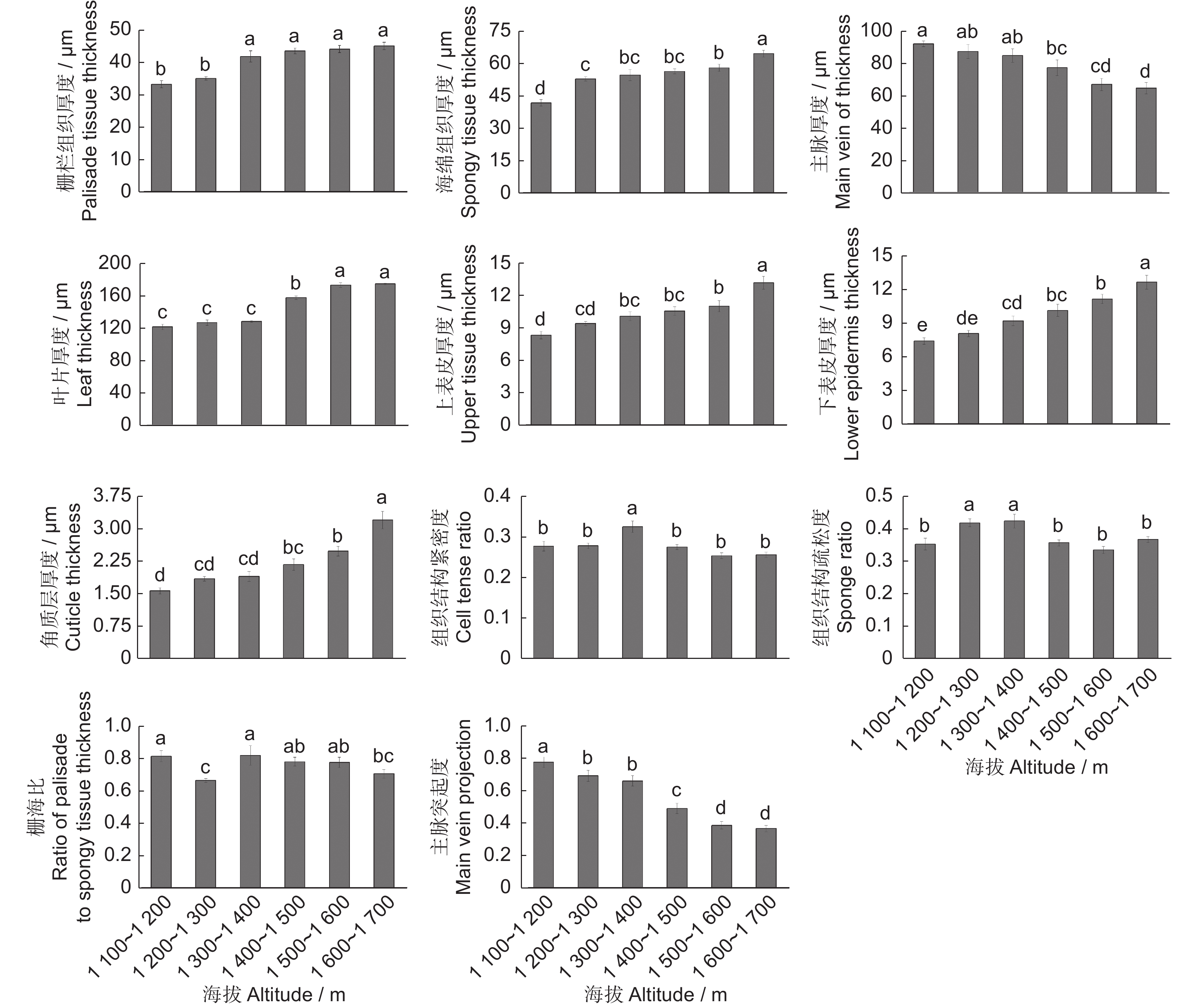
本研究发现,准噶尔山楂叶片栅栏组织厚度、角质层厚度、上下表皮厚度、海绵组织厚度以及叶片厚度均随海拔升高呈上升趋势。其中,从1 200~1 300 m至1 300~1 400 m栅栏组织厚度显著上升;海绵组织厚度在海拔1 100~1 200 m、1 600~1 700 m处与其他海拔差异显著。叶片厚度在海拔1 400~1 500 m时与其他各海拔段差异显著。上下表皮厚度和角质层厚度在1 600~1 700 m时与其他海拔差异显著(图2)。
![]() 图 2 准噶尔山楂叶片解剖结构随海拔梯度的变化趋势不同字母表示不同海拔各参数差异显著(P<0.05)。Figure 2. Variations in leaf anatomical structures in Crataegus songarica with altitudeDifferent letters indicate significant differences in parameters at different altitudes (P<0.05).
图 2 准噶尔山楂叶片解剖结构随海拔梯度的变化趋势不同字母表示不同海拔各参数差异显著(P<0.05)。Figure 2. Variations in leaf anatomical structures in Crataegus songarica with altitudeDifferent letters indicate significant differences in parameters at different altitudes (P<0.05).主脉厚度、主脉突起度随海拔升高呈下降趋势。其中,主脉厚度在1 100~1 200 m时与1 200~1 300、1 300~1 400 m段差异不显著,与其他海拔段差异显著。主脉突起度在1 100~1 200 m段与其他海拔差异显著,1 400~1 500 m段与其他海拔差异显著。组织结构紧密度在1 300~1 400 m处与其他海拔差异显著,其余各海拔间差异不显著。组织结构疏松度在1 200~1 300、1 300~1 400 m处与其他海拔差异显著。栅海比则随海拔升高呈下降-上升-下降的趋势,其中在1 200~1 300 m处显著低于其他海拔区域。
2.4 叶片解剖结构可塑性和相关性分析
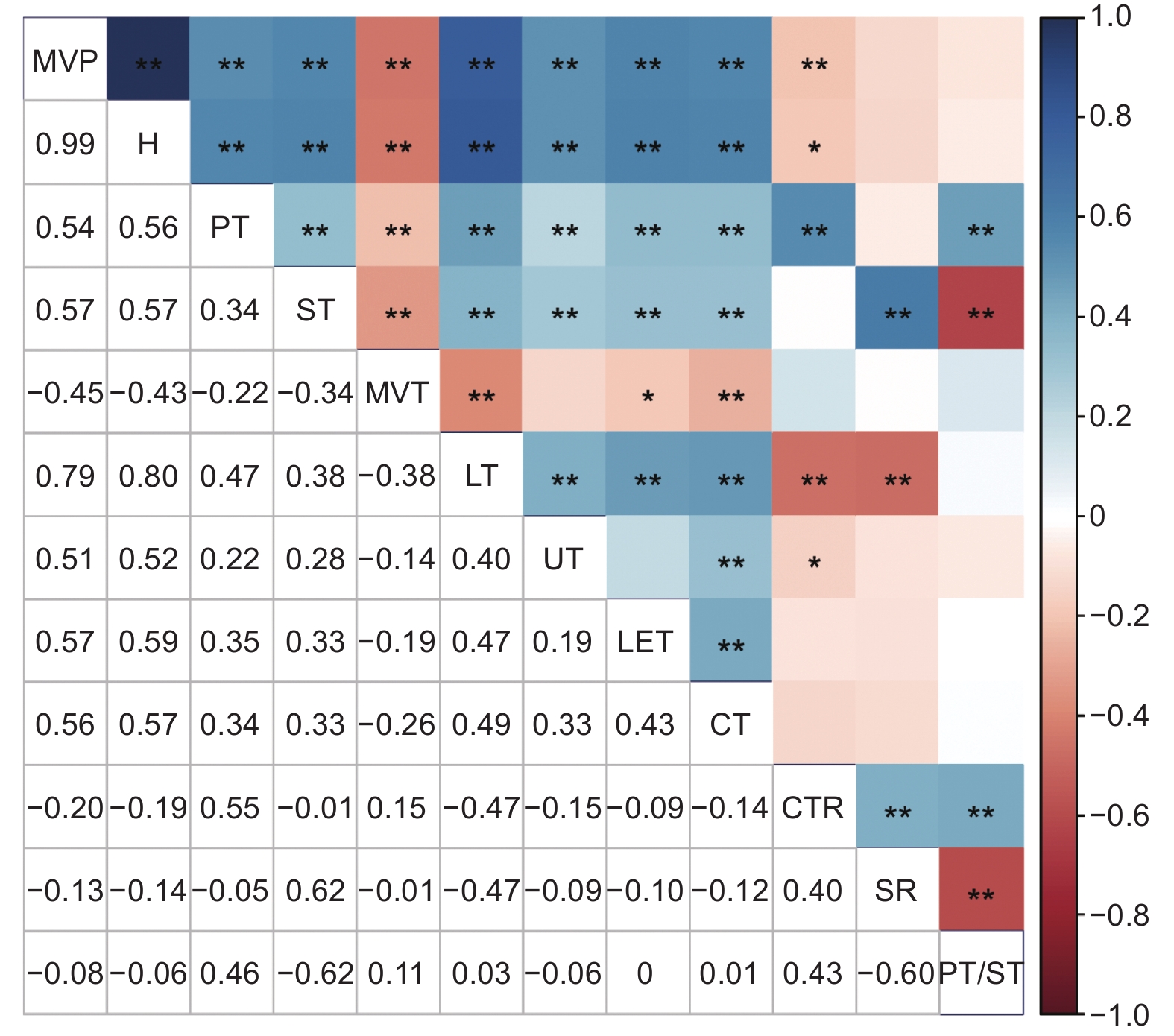
对准噶尔山楂叶片各解剖结构指标进行相关性分析,结果表明,主脉突起度分别与栅栏组织厚度、海绵组织厚度、叶厚、上表皮厚度、下表皮厚度、角质层厚度呈极显著正相关,而与主脉厚度呈极显著负相关。栅栏组织厚度分别与海绵组织厚度、叶厚、上表皮厚度、下表皮厚度、角质层厚度、组织结构紧密度、栅海比呈极显著正相关,而与主脉厚度呈极显著负相关。叶厚分别与上表皮厚度、下表皮厚度、角质层厚度呈极显著正相关,但与组织结构紧密度、组织结构疏松度均呈极显著负相关(图3)。
![]() 图 3 准噶尔山楂叶片解剖结构相关性分析Figure 3. Correlation analysis of leaf anatomical structures in Crataegus songarica
图 3 准噶尔山楂叶片解剖结构相关性分析Figure 3. Correlation analysis of leaf anatomical structures in Crataegus songarica计算准噶尔山楂叶片各项解剖结构指标的可塑性指数,结果发现,所有数值均在0.26~0.49(表3)。其中,主脉突起度可塑性指数最大,其次为角质层厚度,最小为栅栏组织。
表 3 准噶尔山楂叶片解剖结构可塑性指数Table 3. Plasticity index of leaf anatomical structures in Crataegus songarica指标 Index 可塑性指数 PI 指标 Index 可塑性指数 PI 栅栏组织厚度 0.26 角质层厚度 0.45 海绵组织厚度 0.28 叶片组织结构紧密度 0.29 主脉厚度 0.40 叶片组织结构疏松度 0.30 叶片厚度 0.27 栅海比 0.33 上表皮厚度 0.33 主脉突起度 0.49 下表皮厚度 0.38 3. 讨论
3.1 叶片功能性状对海拔的响应
海拔梯度包含了温度、光照等诸多环境因子的变化,有利于研究植物对环境的适应性。在海拔造成的异质环境的长期作用下,植物在功能性状上产生一系列的适应性变化[21]。比叶面积、比叶重可以反映植物的资源周转策略[22]。本研究发现,随海拔的升高,准噶尔山楂的比叶面积逐渐减小,比叶重增大。这可能是由于随海拔的升高,紫外线辐射变强,植物通过减少比叶面积来降低强光辐射对叶片的伤害,同时较高的比叶重可以提高植物在高海拔区域应对逆境胁迫的能力[23]。另一方面,在低海拔区域,植物为了应对激烈的种间竞争,一般通过增加比叶面积来满足自身生长的需求,因此低海拔区域比叶面积通常高于高海拔地区。叶干物质含量不仅体现植物在不同环境下的适应策略[24],也可反映植物结构性碳投资的多少,叶干物质含量越高,表明结构性碳投资越多[25, 26]。本研究中叶干物质含量沿海拔增加,这可能是植物为了适应高海拔区域的恶劣环境,降低水分吸收效率,将自身合成的养分储存到组织器官中[27]。较高的结构性碳投资使得准噶尔山楂可以抵御一定的低温胁迫,从而适应高海拔区域的极端环境。可塑性指数可以揭示植物对环境变化的响应程度[28],本研究结果表明,比叶重和叶干物质含量的可塑性指数较高,说明准噶尔山楂叶片在高海拔区域主要通过调节比叶重、叶干物质含量来适应环境。叶绿素作为植物主要的光合色素,其含量变化会影响植物的光合能力[29],本研究发现,随着海拔的升高,叶绿素含量呈下降趋势,这与胡启鹏等[30]的结论相同。叶绿素的合成通过酶催化进行,而酶活性主要受温度的影响[31]。高海拔区域气温较低,酶活性减弱,叶绿素的合成受到抑制,同时光照强度随着海拔升高而增加,准噶尔山楂通过降低叶片中叶绿素含量来减少过度辐射对叶片的伤害[32],以适应高海拔环境。
综上所述,准噶尔山楂对不同海拔的适应策略不同,可能与叶经济谱有关。叶经济谱的一端是“获取型”资源利用策略,这种策略使得植物往往具有大的比叶面积、低的叶干物质含量、较快的养分获取和利用速率[33],因此有利于其在低海拔区域快速生长,扩大种间竞争优势;另一端是“保守型”资源利用策略,一般表现为叶片养分含量低、相对生长速率慢等,植物通过降低光合、蒸腾速率,储存结构物质来应对外界极端环境的胁迫[34]。利用这种“保守型”策略,准噶尔山楂可以通过增加叶干物质含量、降低比叶面积等措施来适应高海拔地区恶劣的环境。
3.2 叶片解剖结构对海拔的响应
叶片解剖结构体现了植物对环境的适应策略[35]。本研究发现,准噶尔山楂叶片厚度、栅栏组织厚度、海绵组织厚度、角质层厚度等均随海拔的升高而增加。在高海拔区域,生境条件恶劣,风速较强,温度降低,大气的保温作用减弱,较厚的叶片可以避免植物受低温胁迫[36]。叶片厚度增大,一方面可有效降低水分散失速率,增加保水能力,另一方面可有效防止强光辐射对光合系统造成损伤[37]。此外,高海拔区域的栅栏组织和海绵组织比低海拔区域发达。作为叶绿体的主要分布区域,发达的栅栏组织既可以提高准噶尔山楂叶片的光合能力,又有利于增加叶绿体的移动空间,从而减轻因强光对叶肉细胞的损伤[38]。较厚的海绵组织有利于增加光合作用中气体的交换空间[39]。叶片组织结构紧密度越高,植物抵抗低温环境和生理干旱的能力越强[40]。本研究结果显示,在海拔1300~1400 m处准噶尔山楂叶片组织结构紧密度最高,在此海拔区域内,其抗逆性高于其他海拔区域,这是植物对外界环境因素变化的适应结果。这符合“中度干扰假说”,即分布于中间海拔的植物生长优于较低或较高海拔区域[41]。此外,随着海拔的升高,准噶尔山楂叶片主脉厚度逐渐减小。主脉厚度减小,有利于提高叶片吸收、利用光的能力,从而使更多的光辐射能够穿透叶片表皮细胞到达叶肉组织,提高光合能力[42]。可塑性分析结果也显示主脉突起度可塑性最强,说明准噶尔山楂主要通过改变主脉厚度和叶片厚度的比值来适应外界的强紫外线和低温。
3.3 叶片性状间的协同关系
植物叶片性状间联系紧密,随着环境梯度的变化,植物通过协调各叶片性状变化来适应环境[43]。随着海拔的升高,准噶尔山楂叶片功能性状和解剖结构的变化趋势不尽相同,但各性状间存在一定的协同变化。如,叶面积与叶长、比叶面积呈正相关,与比叶重呈负相关,叶厚与栅栏组织厚度、角质层厚度呈正相关。这是因为低海拔地区植株更倾向于自身的生长发育,较大的叶面积和较高的叶绿素含量有利于加快植物的光合速率[44],较窄的栅栏组织、海绵组织以及较薄的角质层则有利于促使叶片快速获取更多的光能,合成营养物质供其自身生长发育。在高海拔区域,准噶尔山楂受低温胁迫影响,更倾向于保存资源,通过增加叶厚、叶干物质含量等来提高自身抗性,并减少叶面积和叶绿素含量来降低蒸腾速率。
4. 结论
准噶尔山楂叶片在不同海拔梯度上的适应策略存在差异,在低海拔主要通过增大叶面积、叶绿素含量等来快速获取生长所需资源,但在高海拔则产生一系列与防御相关的措施(增加叶厚、栅栏组织厚度、海绵组织厚度和角质层厚度以及减小叶面积、叶绿素含量、主脉厚度等)来适应环境中的低温胁迫和强光辐射。
-
![]()
图 1 准噶尔山楂叶片功能性状间相关性分析
Figure 1. Correlation analysis of leaf functional traits in Crataegus songarica
*:P<0.05;**:P<0.01. Same below.
![]()
图 2 准噶尔山楂叶片解剖结构随海拔梯度的变化趋势
不同字母表示不同海拔各参数差异显著(P<0.05)。
Figure 2. Variations in leaf anatomical structures in Crataegus songarica with altitude
Different letters indicate significant differences in parameters at different altitudes (P<0.05).
![]()
图 3 准噶尔山楂叶片解剖结构相关性分析
Figure 3. Correlation analysis of leaf anatomical structures in Crataegus songarica
表 1 样地基本信息
Table 1 Basic information of sample plots
样地号
Sampling site海拔
Altitude / m纬度
Latitude / N经度
Longitude / E坡度
Slope / °1 1 156 44°22′20.19″ 80°57′43.15″ 13.89 2 1 180 44°22′20.08″ 80°57′40.26″ 16.13 3 1 195 44°22′20.23″ 80°57′41.51″ 17.98 4 1 209 44°22′20.22″ 80°57′44.35″ 18.85 5 1 236 44°22′20.08″ 80°57′44.25″ 19.56 6 1 274 44°22′20.94″ 80°57′45.13″ 20.23 7 1 312 44°22′20.57″ 80°57′49.05″ 20.63 8 1 358 44°22′21.26″ 80°57′50.12″ 21.05 9 1 376 44°22′21.27″ 80°57′51.26″ 22.47 10 1 410 44°22′22.28″ 80°57′52.41″ 23.50 11 1 430 44°22′26.43″ 80°58′15.16″ 24.65 12 1 483 44°22′23.30″ 80°57′54.02″ 24.78 13 1 510 44°22′22.31″ 80°57′55.33″ 25.01 14 1 538 44°22′23.32″ 80°57′56.21″ 25.58 15 1 580 44°22′21.29″ 80°57′59.58″ 27.83 16 1 612 44°22′22.80″ 80°57′43.92″ 29.92 17 1 623 44°22′23.59″ 80°57′45.83″ 30.97 18 1 630 44°23′13.15″ 80°57′37.41″ 31.65  下载: 导出CSV
下载: 导出CSV
表 2 不同海拔叶片功能性状变化特征
Table 2 Changes of leaf functional traits at different altitudes
海拔
A / m叶长
LL / cm叶宽
LW / cm叶长宽比
LL/LW叶面积
LA / cm2比叶面积
SLA / cm2/g比叶重
SLW / mg/cm2叶干物质含量
LDMC / g/g叶绿素含量
SPAD1 100~1 200 5.92±0.15a 5.73±0.16a 1.00±0.04c 17.56±0.97a 170.58±10.22a 6.03±0.47c 0.37±0.03c 42.01±1.51a 1 200~1 300 5.77±0.16ab 5.57±0.12a 1.02±0.04bc 16.19±0.81ab 140.34±8.75b 6.08±0.43c 0.46±0.03bc 40.34±0.87a 1 300~1 400 5.68±0.13ab 5.38±0.17ab 1.10±0.05abc 14.56±0.84bc 130.82±7.88bc 6.75±0.38c 0.49±0.04ab 39.73±0.81a 1 400~1 500 5.59±0.18ab 5.38±0.21ab 1.14±0.04ab 13.77±0.61c 118.24±10.37bcd 7.57±1.23c 0.49±0.03ab 39.28±1.33a 1 500~1 600 5.42±0.19ab 4.98±0.17bc 1.16±0.05a 13.25±0.90c 113.81±7.68cd 11.12±1.08b 0.50±0.05ab 38.93±1.15a 1 600~1 700 5.39±0.18b 4.61±0.12c 1.18±0.04a 7.88±0.59d 96.10±6.81d 14.17±0.69a 0.58±0.03a 34.89±0.89b 注:数据显示为平均值±标准误,同列不同字母表示不同海拔各参数差异显著(P<0.05)。 Notes: Data is shown as Mean±Standard error, and different letters in the same column indicate significant differences in parameters at different altitudes (P<0.05).
下载: 导出CSV
表 3 准噶尔山楂叶片解剖结构可塑性指数
Table 3 Plasticity index of leaf anatomical structures in Crataegus songarica
指标 Index 可塑性指数 PI 指标 Index 可塑性指数 PI 栅栏组织厚度 0.26 角质层厚度 0.45 海绵组织厚度 0.28 叶片组织结构紧密度 0.29 主脉厚度 0.40 叶片组织结构疏松度 0.30 叶片厚度 0.27 栅海比 0.33 上表皮厚度 0.33 主脉突起度 0.49 下表皮厚度 0.38
下载: 导出CSV
-
[1] Ma M,Zhao CZ,Kang MP,Li XY,Zhang ZW,Bai XQ. Periploca sepium leaf dry weight,area,and thickness scale differently under different light environments in Taohe riparian forests[J]. Flora,2023,301:152255. doi: 10.1016/j.flora.2023.152255
[2] Xu R,Cheng SD,Zhou J,Tigabu M,Ma XQ,Li M. Intraspecific variations in leaf functional traits of Cunninghamia lanceolata provenances[J]. BMC Plant Biol,2023,23(1):92. doi: 10.1186/s12870-023-04097-y
[3] De Cárcer PS,Vitasse Y,Peñuelas J,Jassey VEJ,Buttler A,Signarbieux C. Vapor-pressure deficit and extreme climatic variables limit tree growth[J]. Glob Chang Biol,2018,24(3):1108−1122. doi: 10.1111/gcb.13973
[4] 黄伟佳,刘春,刘岳,黄斌,李定强,袁再健. 南岭山地不同海拔土壤生态化学计量特征及影响因素[J]. 生态环境学报,2023,32(1):80−89. Huang WJ,Liu C,Liu Y,Huang B,Li DQ,Yuan ZJ. Soil ecological stoichiometry and its influencing factors at different elevations in Nanling Mountains[J]. Ecology and Environmental Sciences,2023,32(1):80−89.
[5] 仲小茹,张露,潘昕昊,叶雄英,黄献峰,等. 常绿阔叶林米槠叶片功能性状对不同海拔梯度的响应与适应[J]. 江西农业大学学报,2022,44(6):1438−1447. Zhong XR,Zhang L,Pan XH,Ye XY,Huang XF,et al. Response and adaptation of leaf functional traits to different altitudes in evergreen broad-leaved forest of Castanopsis carlesii[J]. Acta Agriculturae Universitatis Jiangxiensis,2022,44(6):1438−1447.
[6] Chai YF,Zhang XF,Yue M,Liu X,Li Q,et al. Leaf traits suggest different ecological strategies for two Quercus species along an altitudinal gradient in the Qinling Mountains[J]. J For Res,2015,20(6):501−513. doi: 10.1007/s10310-015-0496-z
[7] 陈艳艳,黄轩,黄晓霞,程小毛. 贡嘎山不同海拔峨眉冷杉叶片生态解剖结构特性研究[J]. 西南林业大学学报,2020,40(6):160−165. Chen YY,Huang X,Huang XX,Cheng XM. An eco-anatomical study on Abies fabri leaves at gradient elevation in Gongga mountain[J]. Journal of Southwest Forestry University,2020,40(6):160−165.
[8] Liu WS,Zheng L,Qi DH. Variation in leaf traits at different altitudes reflects the adaptive strategy of plants to environmental changes[J]. Ecol Evol,2020,10(15):8166−8175. doi: 10.1002/ece3.6519
[9] 郭文文,卓么草,方江平,卢杰,权红,任毅华. 藏东南色季拉山薄毛海绵杜鹃叶解剖结构特征与环境适应性[J]. 西北植物学报,2020,40(5):811−818. Guo WW,Zhuo MC,Fang JP,Lu J,Quan H,Ren YH. Anatomical characteristics and environmental adaptability of Rhododendron aganniphum var. schizopeplum leaf in Sejila mountain,southeastern Tibet[J]. Acta Botanica Boreali-Occidentalia Sinica,2020,40(5):811−818.
[10] 范贺娟,来风兵,曹家睿,陈蜀江,许超宗. 天山野果林区滑坡景观时空演变及生态风险预测[J]. 山地学报,2020,38(2):231−240. Fan HJ,Lai FB,Cao JR,Chen SJ,Xu CZ. Spatio-temporal evolution and ecological risk prediction of landslide landscape in the Tianshan wild fruit forest area[J]. Mountain Research,2020,38(2):231−240.
[11] 盛芳,陈淑英,田嘉,李鹏,秦雪,等. 新疆准噶尔山楂不同居群的遗传多样性[J]. 生物多样性,2017,25(5):518−530. doi: 10.17520/biods.2016369 Sheng F,Chen SY,Tian J,Li P,Qin X,et al. Genetic diversity of Crataegus songorica in Xinjiang[J]. Biodiversity Science,2017,25(5):518−530. doi: 10.17520/biods.2016369
[12] 刘欢,廖康,刘娟,赵世荣,孙琪,曹倩. 新疆野生山楂资源遗传多样性及亲缘关系的ISSR分析[J]. 经济林研究,2016,34(2):19−23. Liu H,Liao K,Liu J,Zhao SR,Sun Q,Cao Q. Analysis of genetic diversity and genetic relationship of wild hawthorn resources in Xinjiang by ISSR markers[J]. Nonwood Forest Research,2016,34(2):19−23.
[13] 刘影,张相锋,赵玉,甘宇平. 新疆濒危野生樱桃李的种群结构与动态[J]. 生态学杂志,2013,32(7):1762−1769. Liu Y,Zhang XF,Zhao Y,Gan YP. Population structure and its dynamics of wild Prunus divaricata in Xinjiang,Northwest China[J]. Chinese Journal of Ecology,2013,32(7):1762−1769.
[14] 侯博,许正. 中国伊犁野生果树及近缘种研究[J]. 干旱区研究,2006,23(3):453−458. Hou B,Xu Z. Study on wild fruit trees and their kindred species in the Yili valley,China[J]. Arid Zone Research,2006,23(3):453−458.
[15] 魏海霞,霍艳玲,周忠科,张治国. 唐古特白刺叶功能性状沿气候梯度的变异特征[J]. 生态学报,2022,42(20):8343−8351. Wei HX,Huo YL,Zhou ZK,Zhang ZG. Variations in leaf traits of Nitraria tangutorum along a climatic gradient[J]. Acta Ecologica Sinica,2022,42(20):8343−8351.
[16] 孙慧兰,李卫红,杨余辉,杨玉海. 伊犁山地不同海拔土壤有机碳的分布[J]. 地理科学,2012,32(5):603−608. Sun HL,Li WH,Yang YH,Yang YH. Soil organic carbon changing with altitudes on the Ili mountainous region[J]. Scientia Geographica Sinica,2012,32(5):603−608.
[17] 李明,胡霞,苗兴军,许正,赵忠. 基于SRAP标记的伊犁河谷野生杏群体遗传多样性研究[J]. 园艺学报,2016,43(10):1980−1988. Li M,Hu X,Miao XJ,Xu Z,Zhao Z. Genetic diversity analysis of wild apricot (Prunus armeniaca) populations in the Ili valley as revealed by SRAP markers[J]. Acta Horticulturae Sinica,2016,43(10):1980−1988.
[18] 羊海军,崔大方,许正,林培钧. 中国天山野果林种子植物组成及资源状况分析[J]. 植物资源与环境学报,2003,12(2):39−45. Yang HJ,Cui DF,Xu Z,Lin PJ. Analysis on the components and resource situation of seed plants in the wild fruit forest in Tianshan Mountain in China[J]. Journal of Plant Resources and Environment,2003,12(2):39−45.
[19] 杨蕾,吕海英,李进,马雪,艾克拜尔·依米提. 新疆天山野果林准噶尔山楂种群结构与动态分析[J]. 西北植物学报,2018,38(12):2314−2323. Yang L,Lü HY,Li J,Ma X,Akbar·Yimit. Structure and dynamic analysis of Crataegus songarica K. Koch population in Tianshan wild fruit forest of Xinjiang[J]. Acta Botanica Boreali-Occidentalia Sinica,2018,38(12):2314−2323.
[20] Gratani L,Covone F,Larcher W. Leaf plasticity in response to light of three evergreen species of the Mediterranean maquis[J]. Trees,2006,20(5):549−558. doi: 10.1007/s00468-006-0070-6
[21] 杨靖,钟乾娟,王崇云,何明超,张天碧,等. 云南药山黄背栎种群不同海拔空间格局分析[J]. 生态科学,2019,38(5):111−118. Yang J,Zhong QJ,Wang CY,He MC,Zhang TB,et al. Spatial pattern analysis of Quercus pannosa population at different altitudes in Yunnan Yaoshan[J]. Ecological Science,2019,38(5):111−118.
[22] 肖燕,张科燕,张树斌,张教林. 羊蹄甲属藤本和树木叶片热值与建成成本的比较研究[J]. 植物科学学报,2020,38(3):428−436. Xiao Y,Zhang KY,Zhang SB,Zhang JL. Differences in leaf caloric values and construction costs between liana and tree species in Bauhinia[J]. Plant Science Journal,2020,38(3):428−436.
[23] 郑梦娜,贾傲,陈之光,廣田充,唐艳鸿,等. 青藏高原矮火绒草(Leontopodium nanum)叶片性状对海拔高度变化的响应[J]. 生态学报,2022,42(24):10305−10316. Zheng MN,Jia A,Chen ZG,Hirota M,Tang YH,et al. Responses of leaf traits of Leontopodium nanum to altitude changes on the Qinghai-Tibetan Plateau[J]. Acta Ecologica Sinica,2022,42(24):10305−10316.
[24] 郑大柽,秦倩倩,邱聪,刘艳红. 重度火烧1年后林下植被的物种多样性及叶功能性状[J]. 应用与环境生物学报,2022,28(6):1601−1607. Zheng DC,Qin QQ,Qiu C,Liu YH. Species diversity and leaf functional traits of understory vegetation one year after severe burning[J]. Chinese Journal of Applied and Environmental Biology,2022,28(6):1601−1607.
[25] Momo ST,Ploton P,Martin-Ducup O,Lehnebach R,Fortunel C,et al. Leveraging signatures of plant functional strategies in wood density profiles of African trees to correct mass estimations from terrestrial laser data[J]. Sci Rep,2020,10(1):2001. doi: 10.1038/s41598-020-58733-w
[26] Bonilla D,Aldana AM,Cárdenas S,Sanchez A. Functional divergence between várzea and igapó forests:a study of functional trait diversity in the Colombian Orinoco basin[J]. Forests,2020,11(11):1172. doi: 10.3390/f11111172
[27] 高风,文仕知,韦铄星,欧汉彪,王智慧,等. 不同种源香合欢种子和叶表型性状变异分析[J]. 中南林业科技大学学报,2023,43(2):57−66. Gao F,Wen SZ,Wei SX,Ou HB,Wang ZH,et al. Variation analysis of seed and leaf phenotypic traits of Albizia odoratissima from different provenances[J]. Journal of Central South University of Forestry & Technology,2023,43(2):57−66.
[28] 王姝,周道玮. 植物表型可塑性研究进展[J]. 生态学报,2017,37(24):8161−8169. Wang S,Zhou DW. Research on phenotypic plasticity in plants:an overview of history,current status,and development trends[J]. Acta Ecologica Sinica,2017,37(24):8161−8169.
[29] Reitz O,Bogena H,Neuwirth B,Sanchez-Azofeifa A,Graf J,et al. Environmental drivers of gross primary productivity and light use efficiency of a temperate spruce forest[J]. J Geophys Res:Biogeosci,2023,128(2):e2022JG007197. doi: 10.1029/2022JG007197
[30] 胡启鹏,郭志华,孙玲玲,王彬. 长白山林线树种岳桦幼树叶功能型性状随海拔梯度的变化[J]. 生态学报,2013,33(12):3594−3601. doi: 10.5846/stxb201205160724 Hu QP,Guo ZH,Sun LL,Wang B. Response of leaf functional traits of Betula ermanii saplings to the altitudinal variation[J]. Acta Ecologica Sinica,2013,33(12):3594−3601. doi: 10.5846/stxb201205160724
[31] 邵文静,张今杰,盖胜男,魏玉磊,李嘉欣,等. 低温胁迫下高粱幼苗叶片生理变化及相关基因表达分析[J]. 农业生物技术学报,2021,29(5):857−870. Shao WJ,Zhang JJ,Ge SN,Wei YL,Li JX,et al. Physiological changes and related genes expression analysis of sorghum (Sorghum bicolor) seedlings under low temperature stress[J]. Journal of Agricultural Biotechnology,2021,29(5):857−870.
[32] 闫景明,周晓兵,张静,陶冶. 新疆野苹果枝条化学计量海拔变异特征研究[J]. 干旱区研究,2021,38(2):450−459. Yan JM,Zhou XB,Zhang J,Tao Y. Variation in one-year-old branch stoichiometry of Malus sieversii at different altitudes and the influencing factors in Tianshan Mountains,China[J]. Arid Zone Research,2021,38(2):450−459.
[33] Reich PB,Hobbie SE,Lee TD. Plant growth enhancement by elevated CO2 eliminated by joint water and nitrogen limitation[J]. Nat Geosci,2014,7(12):920−924. doi: 10.1038/ngeo2284
[34] Wright IJ,Reich PB,Westoby M,Ackerly DD,Baruch Z,et al. The worldwide leaf economics spectrum[J]. Nature,2004,428(6985):821−827. doi: 10.1038/nature02403
[35] Ochirova SS,Dordzhieva VI,Boskhomdzhieva ED,Kolivaev BB. Anatomical structure of the leaf of Filipéndula hexapetala[J]. IOP Conf Ser:Earth Environ Sci,2021,839(5):052014. doi: 10.1088/1755-1315/839/5/052014
[36] 王菲,程小毛,肖云龙,黄晓霞. 千家寨野生古茶树叶片解剖结构和化学组分计量特征对海拔梯度的适应[J]. 生态学杂志,2021,40(7):1958−1968. Wang F,Cheng XM,Xiao YL,Huang XX. Adaptation of leaf anatomical structure and stoichiometric characteristics of wild ancient tea tree to different altitudes in Qianjiazhai[J]. Chinese Journal of Ecology,2021,40(7):1958−1968.
[37] 刘玉萍,周勇辉,吕婷,张晓宇,苏旭. 青藏高原3种野豌豆光合生理特性的比较[J]. 草地学报,2017,25(1):122−129. Liu YP,Zhou YH,Lü T,Zhang XY,Su X. Comparative studies of photosynthetic characteristics of three Vicia species from the Qinghai-Tibet Plateau[J]. Acta Agrestia Sinica,2017,25(1):122−129.
[38] Mendes KR,Marenco RA. Stomatal opening in response to the simultaneous increase in vapor pressure deficit and temperature over a 24-h period under constant light in a tropical rainforest of the central Amazon[J]. Theor Exp Plant Physiol,2017,29(4):187−194. doi: 10.1007/s40626-017-0094-x
[39] Kumari S. Effects of nitrogen levels on anatomy,growth,and chlorophyll content in sunflower (Helianthus annuus L. ) leaves[J]. J Agric Sci,2017,9(8):208−219.
[40] 滕尧,李安定,郝自远,张洪亮,张丽敏,蔡国俊. 西番莲解剖结构特征及低温胁迫下叶片结构与抗寒性的关系[J]. 浙江农业学报,2018,30(11):1849−1858. Teng Y,Li AD,Hao ZY,Zhang HL,Zhang LM,Cai GJ. Anatomical structure of Passiflora caerulea L. and relationship between leaf structure and cold resistance under low temperature stress[J]. Acta Agriculturae Zhejiangensis,2018,30(11):1849−1858.
[41] Kiełtyk P. Variation of vegetative and floral traits in the alpine plant Solidago minuta:evidence for local optimum along an elevational gradient[J]. Alp Bot,2018,128(1):47−57. doi: 10.1007/s00035-017-0197-7
[42] 魏丽萍,田耀华,周会平,岩香甩,龚燕雄,原慧芳. 遮荫处理对不同品种橡胶树叶片解剖特征和生长的影响[J]. 热带作物学报,2019,40(1):45−53. Wei LP,Tian YH,Zhou HP,Yan XS,Gong YX,Yuan HF. The influence of shade treatment on leaf anatomical characteristics and growth of different rubber variaties[J]. Chinese Journal of Tropical Crops,2019,40(1):45−53.
[43] 贾婷,宋武云,关新贤,魏智文,陈涵,等. 湿地松针叶功能性状及其对磷添加的响应[J]. 南京林业大学学报(自然科学版),2021,45(6):65−71. Jia T,Song WY,Guan XX,Wei ZW,Chen H,et al. Responses of needle functional traits of Pinus elliottii to phosphorus addition[J]. Journal of Nanjing Forestry University (Natural Sciences Edition)
,2021,45(6):65−71. [44] 刘晓娟,马克平. 植物功能性状研究进展[J]. 中国科学:生命科学,2015,45(4):325−339. doi: 10.1360/N052014-00244 Liu XX,Ma KP. Plant functional traits:concepts,applications and future directions[J]. Scientia Sinica Vitae,2015,45(4):325−339. doi: 10.1360/N052014-00244
-
期刊类型引用(4)
1. 黄文静,郭有艳,喻阳华,罗红,杨广广,杨顺松. 喀斯特区特有植物顶坛花椒不同类型幼苗叶片功能性状. 贵州农业科学. 2025(02): 83-92 .  百度学术
百度学术
2. 冯晓宇,吕世琪,张芸香. 华北落叶松不同海拔更新苗对移栽地海拔的适应性差异. 广西林业科学. 2025(01): 44-51 . 百度学术
3. 刘仁,温黎彬,贺小林,蔡宗明,朱琪,郭志强,魏敏,刘儒. 不同山桐子家系叶片功能性状及养分特征差异. 东北林业大学学报. 2024(08): 60-68 . 百度学术
4. 宁朋,袁治国,乔秋芬,王玲. 新疆乌什县不同海拔黄花软紫草生长及生理特性. 西北植物学报. 2024(09): 1386-1394 . 百度学术
其他类型引用(0)






计量
- 文章访问数: 342
- HTML全文浏览量: 46
- PDF下载量: 195
- 被引次数: 4
