
Prediction of potential distribution of Cupressus gigantea W. C. Cheng & L. K. Fu in China based on optimized MaxEnt modeling
-
摘要:
巨柏(Cupressus gigantea W. C. Cheng & L. K. Fu)是西藏特有种,国家一级保护植物。本研究基于R软件“kuenm”包优化模型参数,建立最优MaxEnt模型,预测了全国范围内巨柏的潜在适宜分布区,并结合贡献率和折刀测验分析了影响其分布的主导环境变量。结果显示,优化模型的表现效果较好,相比默认参数模型,降低了模型的复杂度,提高了模拟预测精度。与河流的距离、温度季节性变化标准差、等温性、坡度、最冷季度降水量是影响巨柏分布的主导环境变量。巨柏在我国的潜在适宜分布区集中在西藏和四川,西藏的林芝、山南、拉萨和日喀则占据了全部中适宜区(2 298 km2)和高适宜区(746 km2),其中高适宜区只分布在林芝(563 km2)和山南(183 km2),约占西藏总面积的0.062%。朗县-米林市的高适宜区为巨柏潜在生境分布的核心地段,建议于此区域优先开展巨柏保护工作。
Abstract:Cupressus gigantea W. C. Cheng & L. K. Fu, a tree species endemic to Tibet and classified as a national first-class protected species in China, requires clear delineation of its potential suitable distribution for effective conservation, introduction, and breeding programs. This study established the MaxEnt model, using the “kuenm” R package to optimize model parameters, to predict the potential distribution of C. gigantea across China. Environmental variables influencing distribution were analyzed through percent contribution and Jackknife tests. The optimized model demonstrated superior predictive accuracy and reduced complexity compared to the default parameter model. Key factors influencing the distribution of C. gigantea included distance from rivers, temperature seasonality, isothermality, slope, and precipitation during the coldest quarter. Predicted suitable distribution areas were primarily located in Tibet and Sichuan, with the cities of Nyingchi, Lhoka, Lhasa, and Rikaze in Tibet encompassing the entirety of moderately suitable (2 298 km2) and highly suitable areas (746 km2). The high suitability area was exclusively distributed in Nyingchi (563 km2) and Lhoka (183 km2), representing about 0.062% of the total area of Tibet. The corridor between Lang County and Milin County emerged as the core habitat for C. gigantea, underscoring the need to prioritize conservation efforts in this critical region.
-
Keywords:
- Cupressus gigantea /
- Habitat /
- Potential distribution /
- MaxEnt /
- Tibet
-
巨柏(Cupressus gigantea W. C. Cheng & L. K. Fu)又名雅鲁藏布江柏木,系柏科柏木属(Cupressus)高大常绿乔木,是我国西藏特有的珍稀树种之一,国家一级保护植物[1]。其现代自然地理分布狭窄,仅见于雅鲁藏布江中游沿江的部分河谷坡地及其支流尼洋河的下游地区,垂直分布范围为海拔2 500~3 400m[2]。现代分布区可分为朗县、米林市和巴宜区(林芝县)3个居群,朗县居群沿雅鲁藏布江呈“走廊式”带状分布,其余两个居群成“岛状”分布[3, 4]。巨柏起源古老,树龄可高达2 500年以上,树形优美,兼具生态、经济、文化价值,对研究柏科植物的系统发育、藏东南气候变化规律、植被演替及其与环境的关系等具有重要意义[5]。受恶劣生存环境、人类活动干扰、种子质量低下以及幼苗定居至幼树建成过程困难等多重因素的影响,巨柏的自然更新面临挑战[4, 6]。因此,探究巨柏适宜生境并加强其保护工作显得尤为迫切。
研究物种的分布格局及其影响因子,制定针对性的物种保护策略,对种质资源保存以及引种保育具有十分重要的意义。生态位模型(Ecological niche models,ENMs)是一种基于生态位理论,通过一定的算法分析物种的分布数据与环境之间的关系,推算满足物种的生态位需求,从而预测物种潜在生境的方法[7]。目前常用的生态位模型有基于遗传算法的规则组合模型(GARP)、生物气候分析和预测系统模型(BIOCLIM)、最大熵模型(MaxEnt)等。其中,MaxEnt模型是基于最大熵理论,寻找最大熵的概率分布来估计物种的概率分布[8, 9]。因其具有性能稳定、准确率较高、操作简单、样本需求量较小等优点,在生态位模型中表现更突出[10, 11],已被广泛应用于珍稀濒危物种的栖息地适宜性评价和潜在分布区评估[12, 13]。
目前对巨柏的研究主要集中在遗传特征[14]、生物学特性[15]、群落生态[16]、扩繁技术[17]、濒危机制和保护[6, 18]等方面。关于巨柏适生区的预测鲜有报道,姚淇等[19]基于MaxEnt模型结合气候、土壤和地形变量预测了巨柏在青藏高原尺度上的适生区分布,但未考虑水文、人类活动等因素的影响,且其构建的模型采用默认软件参数,未进行模型参数优化。目前国内(如甘肃、北京)已有一些巨柏成功种植的相关报道[4],排除人为养护因素的影响,自然分布区之外的其他生境条件是否适宜于巨柏的生存还有待确认。因此,本研究结合野外实地调查和前人的相关研究成果,基于MaxEnt模型和“kuenm”软件包优化模型参数,以期解决两个问题:(1)全国范围内巨柏潜在适宜生境的分布位置和面积如何?(2)影响巨柏分布的主要环境因素是什么?
1. 材料与方法
1.1 巨柏分布数据的获取与处理
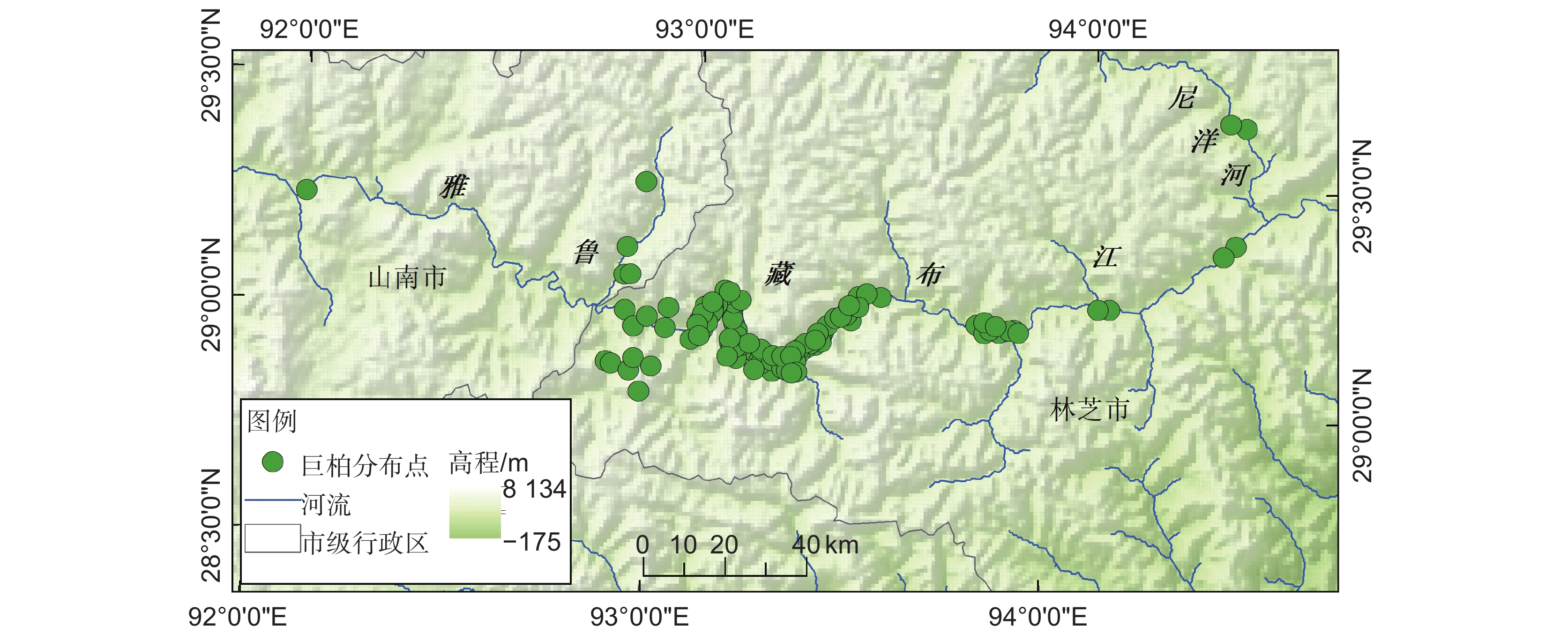
本研究所用的巨柏位点分布数据主要来源于高分辨率遥感影像的目视解译和野外实地调查(2022年7月和2023年5月),同时查阅中国数字植物标本馆(CVH,https://www.cvh.ac.cn/)、中国国家标本资源平台(NSII,http://www.nsii.org.cn/)、全球生物多样性信息数据库网络(GBIF,https://www.gbif.org),并结合文献资料[19]进行补充。为匹配环境数据和减少分布点的空间自相关影响,剔除人工引种栽培的分布点、经纬度精度小于0.01和重复点后,利用ArcGIS10.8软件中的SDMTools工具[20],在直径为1 km的网格内仅保留一个巨柏分布点,共获得123个样本点(图1),导出其经纬度,保存为模型所需的.csv格式。
![]() 图 1 筛选后的巨柏样本点分布图基于自然资源部标准地图服务系统审图号GS(2024)0650号标准地图制作,底图无修改。下同Figure 1. Distribution map of selected sample points of Cupressus giganteaThe map was based on the standard map with review number GS (2024) 0650 downloaded from the standard map service system of the Ministry of Natural Resources, and the base map was not modified. Same below.
图 1 筛选后的巨柏样本点分布图基于自然资源部标准地图服务系统审图号GS(2024)0650号标准地图制作,底图无修改。下同Figure 1. Distribution map of selected sample points of Cupressus giganteaThe map was based on the standard map with review number GS (2024) 0650 downloaded from the standard map service system of the Ministry of Natural Resources, and the base map was not modified. Same below.1.2 环境数据筛选与处理
输入模型的环境变量会直接影响模型预测的结果,筛选对物种分布影响较大的环境变量是准确预测的重要环节[21]。依据文献[22, 23],结合巨柏的生态习性,本研究从气候、地形、水文、土壤、人类活动、植被等6个方面考虑影响其适宜分布的主导环境变量(表1)。具体数据来源如下。(1)气候变量,来源于Worldclim2.1数据库,包括19个生物气候变量和1个基于月均值的太阳辐射变量,空间分辨率为30″。(2)地形变量DEM,取自中国科学院资源环境科学与数据中心(http://www.resdc.cn/),并利用ArcGIS提取坡度和坡向,分辨率为1 km。(3)水文变量,与河流的距离是基于HydroSHEDS数据[24],利用ArcGIS筛选流量 ≤ 5级的中国河流,提取其最邻近欧氏距离。(4)土壤含水量,源自SoilGirds中的高分辨率全球土壤水文属性地图数据集(https://dataverse.harvard.edu/dataset.xhtml?persistentId=doi:10.7910/DVN/UI5LCE),采用田间持水量(Field capacity mean)作为土壤水分变量。为避免MaxEnt模型过度强调分类变量的重要性[25],本文采用砂粒、粉粒、黏粒含量的连续变量替代土壤类型分类,该数据集来源于中国科学院资源环境科学与数据中心。其他土壤变量[26]来源于国家地球系统科学数据中心-土壤分中心(http://soil.geodata.cn),覆盖[0,5)、[5,15)、[15,30)、[30,60)、[60,100)和[100,200) cm 6个土层。鉴于巨柏根系主要分布在0~60 cm的土层[27],本研究仅考虑前4层。土壤变量标注代码为变量简称加土层厚度,如:bd05表示0~5 cm土层的土壤容重。(5) 人类足迹指数(Human footprint index),源自国际地球科学信息网络中心(CIESIN)发布的数据集(https://sedac.ciesin.columbia.edu/data/set/wildareas-v3-2009-human-footprint),分辨率约为1 km,综合建筑环境、人口密度等8个指标,反映了全球人类活动对环境的累积压力,数值越高,表示人类活动强度越大[28]。(6)归一化植被指数(Normalized difference vegetation index,NDVI),源自中国科学院资源环境科学与数据中心,NDVI可指示植被覆盖度,一定程度上能反映物种间的相互作用。本研究采用1998-2018年间的NDVI平均值作为植被变量,分辨率为1 km。
表 1 本研究所用的环境变量Table 1. Environmental variables used in this study类型
Type代码
Code描述
Description单位
Unit说明
Expression气候 bio1 年平均温度 ℃ 等温性bio3=(bio2 / bio7)×100,温度的年较差bio7=bio5−bio6,反映温差特点;温度季节性bio4=温度标准差×100,使用温度的标准偏差表示,反映平均温度的变化幅度;降水季节性bio15采用降水的变异系数表示,反映降水量的季节性分布。
温度和降水通常是影响植物生长发育、繁殖扩散的重要因子。巨柏具有喜温、喜光、趋湿、抗旱、抗寒等生态特性[32]。bio2 每月最高温与最低温差值的平均值 ℃ bio3 等温性 % bio4 温度季节性 − bio5 最热月的最高温度 ℃ bio6 最冷月的最低温度 ℃ bio7 温度的年较差 ℃ bio8 最湿季度的平均温度 ℃ bio9 最干季度的平均温度 ℃ bio10 最热季度的平均温度 ℃ bio11 最冷季度的平均温度 ℃ bio12 年降水量 mm bio13 最湿月的降水 mm bio14 最干月的降水量 mm bio15 降水量季节性变异系数 − bio16 最湿季度的降水量 mm bio17 最干季度的降水量 mm bio18 最热季度的降水量 mm bio19 最冷季度的降水量 mm srad 月均太阳辐射 kJ·m−2·d−1 dem 海拔 m 地形 slope 坡度 ° 地形通过对水热条件的调节和再分配影响物种的组成与分布[33]。 aspect 坡向 − 土壤 thickness 土壤厚度 cm 植物的生息与土壤的结构和理化性质紧密相关。相关研究表明土壤质地、养分、密度、含水量等是影响巨柏群落分布的主要因子[16]。 bd 土壤容重 g/cm3 ph 土壤pH值 − fcm 田间持水量 cm3/cm3 soc 土壤有机碳含量 g/kg tn 土壤全氮含量 g/kg clay 土壤黏粒含量 % slit 土壤粉粒含量 % sand 土壤砂粒含量 % 水文 dr 与河流的距离 m 水线地类是巨柏种子萌发、幼苗幼树构建的重要地类[6]。研究认为水或水中动物可能是巨柏沿雅鲁藏布江分布传播的重要媒介[4]。 人类活动 hf 人类足迹指数 − 伐木、樵采、祭祀拜佛等人类活动是影响巨柏分布的重要因子[4]。 植被 NDVI 归一化植被指数 − 巨柏的更新状况会受灌木层郁闭度和竞争程度的影响[6, 34]。 将上述变量的栅格图层在ArcGIS软件中进行计算、掩膜、重采样、坐标转换等预处理,统一分辨率为1 km,投影坐标为WGS_1984_Albers,应用GIS工具包Circuitscape(https://circuitscape.org/)批量导出为.asc格式备用。环境变量间的高相关性可能导致模型过拟合,影响模型结果的准确性[29]。参考前人文献[30, 31],使用MaxEnt V3.4.4软件默认参数,通过自举法(Bootstrap)重复10次构建模型,并执行折刀测验(Jackknife)评估环境变量的重要性,基于模型的平均结果,排除贡献率小于0的变量(附表1
1 ))。在ArcGIS软件中运用SDMTool工具中的Correlations and Summary Stats工具评估变量的相关性(附图12 )),剔除相关系数|r| >0.8的变量,并在相关性高的变量中保留贡献率和置换重要性较高的变量,最后共得到16个候选环境变量。它们分别为:每月最高温与最低温差值的平均值(bio2)、等温性(bio3)、温度季节性(bio4)、最冷季度的平均温度(bio11)、降水季节性(bio15)、最冷季度的降水量(bio19)、月均太阳辐射(srad)、与河流的距离(dr)、坡向(slope)、海拔(dem)、田间持水量(fcm)、[5,15)cm土壤容重(bd515)、[0,5)cm土壤pH值(ph05)、土壤粉粒含量(slit)、[0,5)cm土壤总氮含量(tn05)以及人类足迹指数(hf)。1.3 模型的建立、优化与评价
本研究利用ArcGIS的子集工具将物种的分布数据随机分为70%训练集和30%测试集。在R 4.2.2软件中通过“kuenm”包(https://github.com/marlonecobos/kuenm)对MaxEnt模型的调控倍率(Regularization multiplier,RM)和特征组合(Feature combination,FC)参数进行优化,该包支持自定义参数组合,以构建候选模型[35]。具体参数设置为:RM从0.5至6,步长为0.5,共12种;FC包括线性(Linear,L)、二次型(Quadratic,Q)、片段化(Hinge,H)、乘积型(Product,P)和阈值性(Threshold,T)5种特征的31种组合[36]。以delta AICc值为模型筛选的标准,该值基于修正的Akaike信息量准则(Akaike information criterion correction,AICc),用于评估模型的预测能力、拟合度和复杂程度。通常将delta AICc<2的模型视为较可靠,选择 delta AICc最小的模型参数为最优设置,以构建最终的MaxEnt模型[35, 37]。此外,设置重复执行10次自举法,以最大训练灵敏度与特异性之和(Maximum training sensitivity plus specificity,MTSS)作为阈值,输出预测结果形式为cloglog[31],执行刀切法并构建响应曲线,其他参数保持默认。利用受试者工作特征曲线(Receiver operating characteristic curve,ROC)下的面积AUC值(Area under the curve,AUC)评估模型的准确性。AUC值的范围为[0,1],该值可反映模型的判断能力,一般认为,当0.8≤AUC<0.9时,预测表现良好;当 0.9≤AUC<1时,预测表现优秀[38, 39]。
1.4 巨柏适生区划分
将模型重复10次得到的平均预测结果导入ArcGIS中进行可视化和适宜区划分。每个像元的阈值为[0,1],这个值代表该物种对该地区环境的相对适应性,值越高表示适应性越强。参照以往研究[37, 40],以MTSS作为适宜性级别划分最低阈值,参照IPCC[41]对于物种存在可能性的定义将巨柏生境适宜区划分为4级:非适宜区(P≤ MTSS)、低适宜区(MTSS<P≤0.33)、中适宜区(0.33<P≤0.66)和高适宜区(P>0.66)。
2. 结果与分析
2.1 模型评价
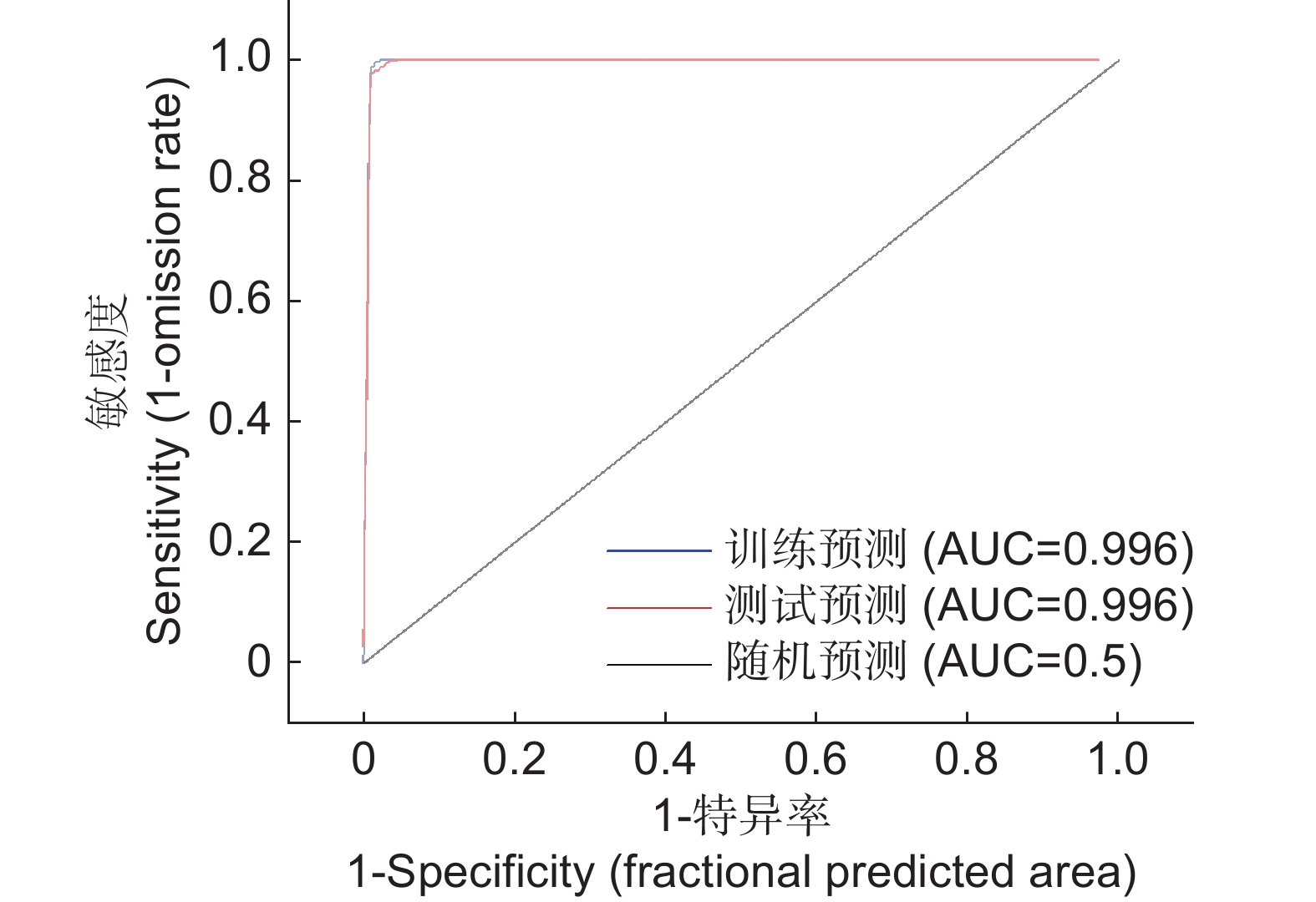
利用“kuenm”软件包对分布数据和环境变量建模,共产生了372个模型(12种调控倍率×31种特征组合)。这些模型均具有统计显著性,其中两个模型满足Akaike信息量准则。基于delta AICc值及其他模型评价指标,本研究确定了最优模型参数配置为:调控倍率RM为1,特征组合FC为“QT”。该模型遗漏率较低(表2),且与默认参数模型相比,其复杂度较低。使用最优参数构建MaxEnt模型,重复运算10次后,结果显示,其平均训练AUC值和测试AUC值均超过0.9,标准差为0.000,表明该模型的拟合程度相对较高,预测效果较好,结果可信(图2)。
表 2 MaxEnt模型“kuenm”包优化评价指标及最优模型参数配置Table 2. Evaluation metrics of MaxEnt model generated by kuenm and optimal model parameter configuration模型
Model平均AUC比值
Mean_AUC_ratiopROC统计显著性
pval_pROC10%遗漏率
Omission_rate_at_10%AICc差值
delta_AICc参数数量
num_parametersM_1_F_QT * 1.999 0 0.216 0.000 22 M_1_F_T 1.999 0 0.297 0.933 18 M_1_F_LQPTH 1.999 0 0.189 55.277 38 注:加粗字体表示“kuenm”包筛选的优化模型,未加粗字体表示默认参数模型; * 指本研究确定的最优模型。 Notes: Bold entries represent optimized model parameters selected by “kuenm” package; non-bold entry indicates default parameter model; * signifies optimal model identified in this study. ![]() 图 2 最终模型的受试者工作曲线(ROC)验证Figure 2. Receiver operating curve (ROC) validation of final model
图 2 最终模型的受试者工作曲线(ROC)验证Figure 2. Receiver operating curve (ROC) validation of final model2.2 环境变量评价
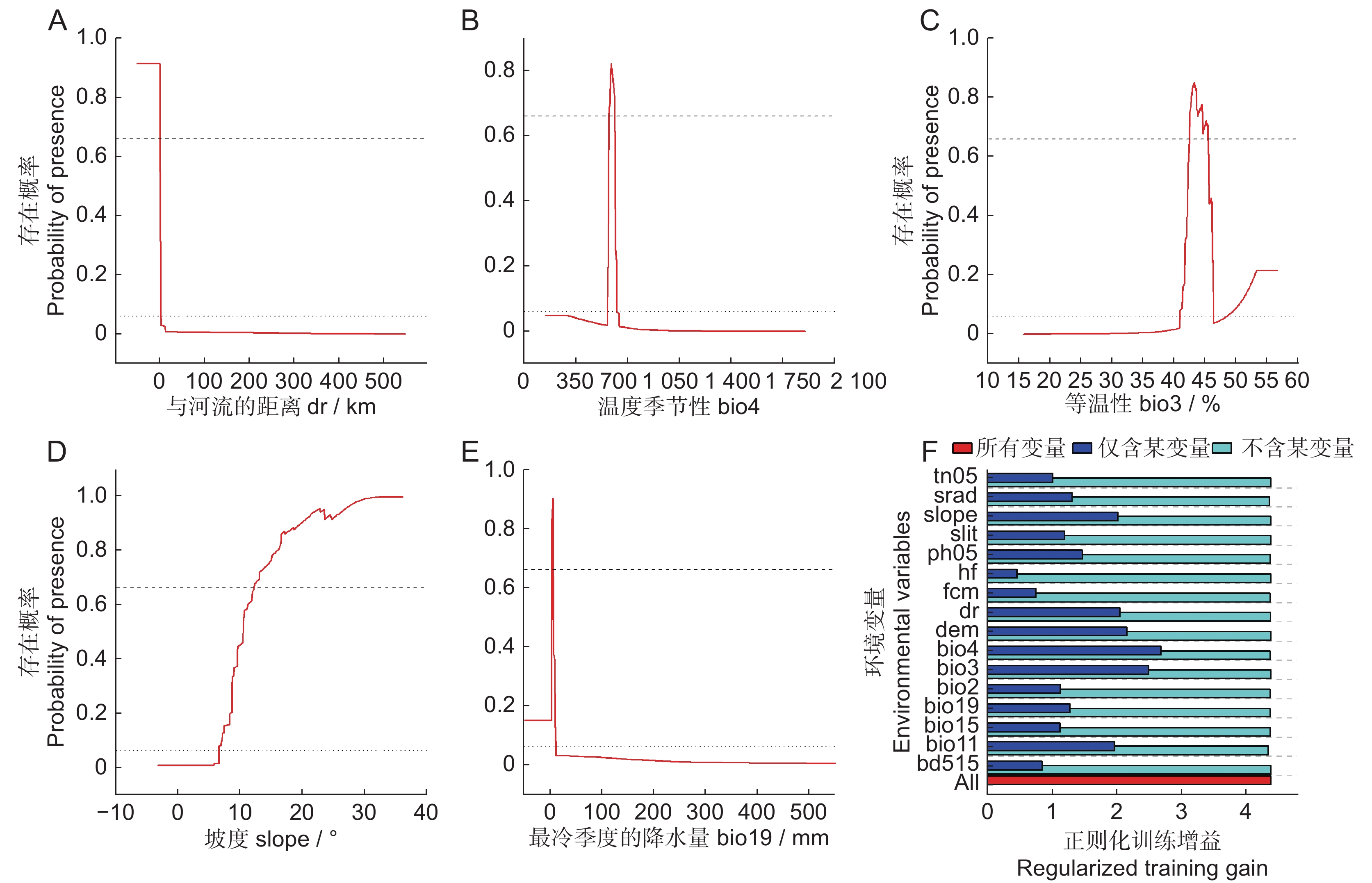
由模型运行的主要环境变量贡献率和置换重要性结果可知(表3),各变量的贡献率差异较小,在诸多变量中贡献率排名前5的变量分别为:与河流的距离、温度季节性、等温性、坡度和最冷季度的降水量,占总贡献率的80.9%。在置换重要性排名中,温度季节性和最冷季度的平均温度对模型的重要性最高,两者的累积重要性高达92.15%。
表 3 主要环境变量的贡献率及重要性分析Table 3. Analysis of contribution rate and importance of main environmental variables环境变量
Environmental variables描述
Description贡献率
Percent contribution / %置换重要性
Permutation importancedr 与河流的距离 19.47 0.07 bio4 温度季节性 18.42 62.16 bio3 等温性 18.23 0.00 slope 坡度 14.11 0.10 bio19 最冷季度的降水量 10.67 0.85 srad 月均太阳辐射 5.68 1.95 dem 海拔 4.02 0.11 bio15 降水季节性 3.14 0.82 fcm 田间持水量 3.05 0.68 ph05 0~5 cm土层的pH值 1.54 2.38 bio11 最冷季度的平均温度 0.94 29.99 依据训练折刀测验结果(图3:F),当模型仅使用单一变量运行时,增益最高的变量是温度季节性,然后依次是等温性、海拔、与河流的距离、坡度和最冷季度的平均温度,表明这些变量对巨柏的潜在适宜区预测模型的解释能力较强。而当模型仅去除某个环境变量时,模型增益变化均比较小。综上所述,气候对巨柏的潜在分布影响最大,其次是水文、地形和土壤。
![]() 图 3 主导环境变量的响应曲线示意图(A~E)以及影响巨柏潜在分布的各环境变量的折刀测验重要性(F)Figure 3. Response curve diagram of dominant environmental variables (A–E) and importance of environmental variables affecting potential distribution of Cupressus gigantea based on Jackknife test (F)
图 3 主导环境变量的响应曲线示意图(A~E)以及影响巨柏潜在分布的各环境变量的折刀测验重要性(F)Figure 3. Response curve diagram of dominant environmental variables (A–E) and importance of environmental variables affecting potential distribution of Cupressus gigantea based on Jackknife test (F)本研究使用单一变量(dr、bio4、bio3、slope和bio19)绘制响应曲线,明确主导环境变量与巨柏存在概率的关系(图3:A~E)。结果发现,存在概率随着与河流的距离增加而迅速下降,而随着坡度的增加呈上升趋势,在32°左右达到最大;与季节性温度变异、等温性和最冷季度降水量之间的关系总体上均表现出先上升后下降的趋势。分别以存在概率P=0.06和P=0.66为阈值,划分巨柏生态适宜性环境变量的范围。结果表明,巨柏的适宜区环境特征为:与河流的距离为0~3 000 m,最适距离区间为0~1 200 m;温度季节性为561~625,最适区间为570~610;等温性为41%~46%和48%~53%,最适等温性范围为42%~45%;坡度为6°~32°,最适坡度范围为12°~32°;最冷季度降水量为0~11 mm,最适范围为3~5 mm。综上,中等偏高水平的温度季节性、中等偏低水平的等温性、较少的最冷季度降水量、与河流距离较近的缓坡和斜坡更有利于巨柏的生存。
2.3 巨柏在中国的潜在生态适宜分布区
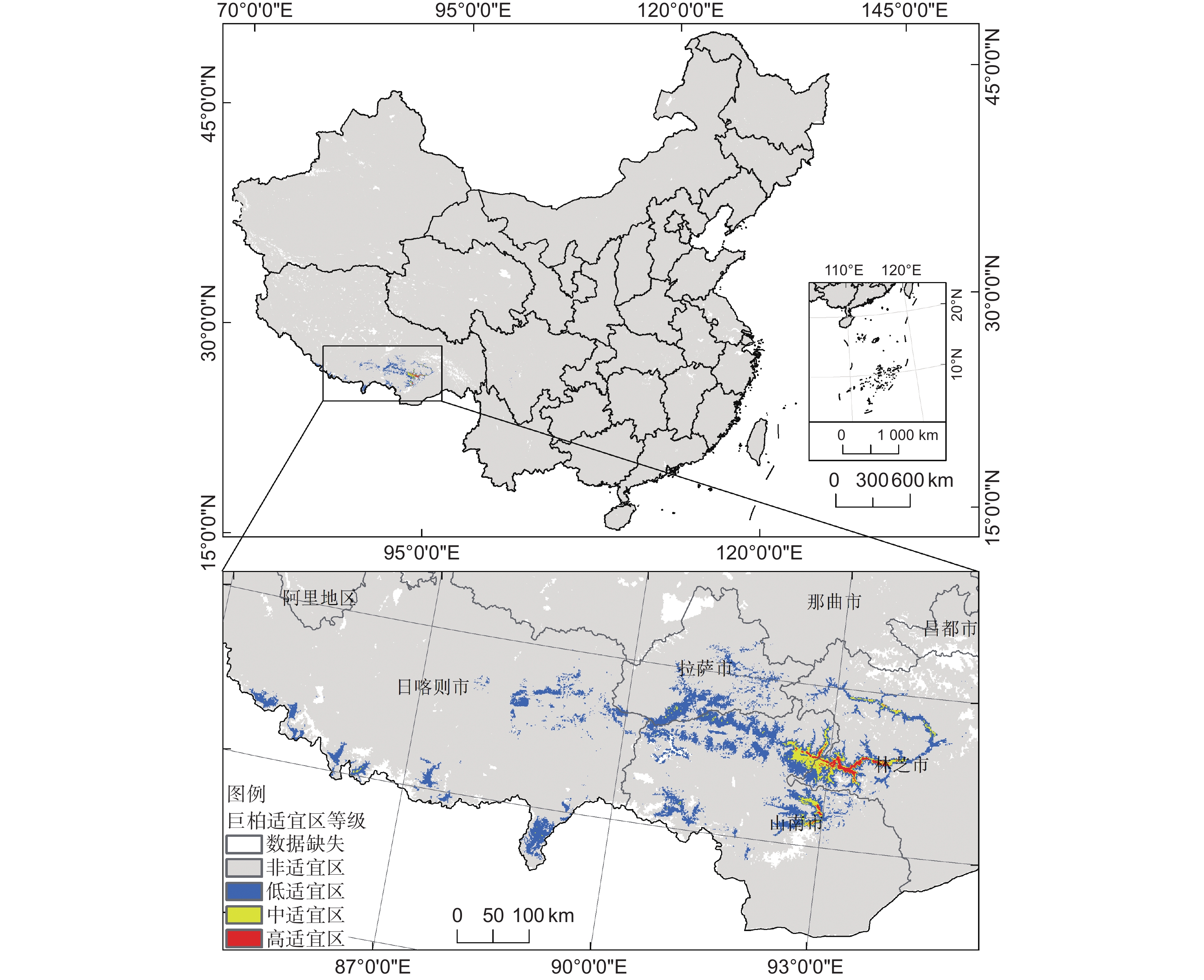
巨柏在全国范围内的高、中生态适宜区均分布在西藏自治区,在四川省分布有低适宜区,其余地方均为非适宜区。巨柏的中、高适宜区集中分布在西藏林芝市的朗县、米林县、巴宜区和工布江达县,面积合计563 km2,以及山南市的加查县、隆子县、桑日县,面积合计183 km2;拉萨市和日喀则市零星分布着中适宜区,面积达62 km2。总体上,适宜巨柏生存的区域主要集中在其现代分布区——雅鲁藏布江中下游和尼洋河沿岸地带,这与目前巨柏的分布区较为一致(表4,图4)。
表 4 全国巨柏主要潜在适宜分布区面积统计Table 4. Statistical details regarding potential suitable distribution areas for C. gigantea in China行政区等级
Administrative
region位置
Location非适宜区
Unsuitable zone / km2低适宜区
Lowly suitable zone / km2中适宜区
Moderately suitable zone / km2高适宜区
Highly suitable zone / km2省级 西藏自治区 1 127 185.00 20 574.00 2 298.00 746.00 四川省 484 766.00 67.00 − − 市级 林芝市 98 206.00 3 629.00 1 116.00 563.00 山南市 65 159.00 8 675.00 1 120.00 183.00 拉萨市 25 356.00 3 234.00 34.00 − 日喀则市 167 216.00 5 031.00 28.00 − 甘孜藏族自治州 148 495.00 65.00 − − 那曲市 340 058.00 5.00 − − 凉山彝族自治州 60 227.00 2.00 − − ![]() 图 4 巨柏在我国的潜在适宜区分布Figure 4. Distribution of potential suitable habitats for Cupressus gigantea in China
图 4 巨柏在我国的潜在适宜区分布Figure 4. Distribution of potential suitable habitats for Cupressus gigantea in China3. 讨论
目前关于主导环境变量的确定缺乏统一标准,有学者采用变量的累计贡献率达到一定程度来确定主导环境变量的数量[37],也有学者选取贡献百分比,如认为贡献率超过10%的变量起主导作用[42]。本研究以10%的贡献率作为参照线确认主导环境变量,其累计贡献率也达到80%。环境变量分析结果表明,巨柏的潜在适宜性分布主要是气候、水文和地形等变量共同作用的结果。中等偏高水平的温度季节性和中等偏低的等温性气候环境较为适宜巨柏生存,这两个气候变量均可反映地区温差的变化幅度,说明巨柏对温差变化的适应性较强。Tang等[43]发现,在中国青藏高原地区,模拟的孑遗植物适生区的等温性大致处于中等水平,与本研究结果相吻合。本研究还发现,巨柏对最冷季度降水量的需求较低,且其存在概率随着降水量的增加先急剧上升而后下降。这可能是巨柏在长期进化中形成的对寒冷干旱环境适应策略的一种表现,与其抗旱抗寒的生物学特性相吻合。另外,本研究结果显示最冷季度平均温度的置换重要性和训练增益排名较靠前,说明该气候因子在预测巨柏分布时的影响也不容忽视。
水文变量与河流的距离是影响巨柏潜在适宜生境分布的关键变量,与前人的观点较一致[4, 44]。水线岸边作为巨柏自然更新的主要阵地,在一定程度上影响着其分布格局[6, 45]。边喜丽等[27]对根系结构特征的研究也发现,生活在水线岸边的巨柏根系具有较明显的向水性,并指出河岸阶地和坡麓地是巨柏的理想迁移保护栽植点。此外,坡度可通过调节区域的降水分布、改变太阳光的入射角度以及影响土壤渗透率等途径塑造不同的生境微气候,也是影响巨柏分布的主要环境因子之一。
本研究预测的巨柏潜在高生态适宜区分布与姚祺等[19]模拟的青藏高原巨柏的潜在高适生区分布区域较为一致,但预测面积相对较小。综合多方面的数据建模和模型参数优化,本研究的预测结果相对而言更为保守,更贴近巨柏的实际潜在适宜分布范围。林芝市的雅鲁藏布江及尼洋河沿岸是巨柏当前主要的天然分布区,也是模型预测的高适宜区集中地带。尤其是米林至朗县一带,中、高适宜区面积大且连续性较强,说明这些区域是巨柏种群生长繁殖的关键区域,未来应以米林-朗县区域的巨柏种群保护为核心。巴宜区的中高适宜区分布少且破碎,针对该区域的保护策略应聚焦于现有种群的生境保护,并构建生态廊道,以促进其与周边种群的基因交流,进而增强种群的稳定性,并扩展其繁殖范围。山南市的加查县有巨柏分布的历史记录[46],隆子县的高适宜区分布在雅鲁藏布江支流西巴霞曲沿岸,该区域尚未有巨柏分布的报道,建议在此开展巨柏引种栽培试点工作,探究其生态适应性及其与当地生态环境之间的关系,以促进巨柏种群的恢复或开辟新的分布区域。
4. 结论
本研究基于优化的MaxEnt模型预测分析,发现巨柏在我国的潜在适宜分布区主要位于西藏自治区和四川省,总面积为23 685 km2。其中,绝大部分中度和高度适宜区分布在西藏的林芝市和山南市,面积为2 982 km2,可以优先考虑在该区域开展巨柏的保护和推广种植工作。
水文、气候和地形因素是影响巨柏潜在适宜生境分布的主导环境变量。巨柏的最适宜生境特征为:与河流的距离为0~1 200 m,季节性温度变异为570~610,等温性为42%~45%,坡度为12°~32°,最冷季度降水量为3~5 mm。
1 如需查阅附表内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。2 如需查阅附图内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。 -
![]()
图 1 筛选后的巨柏样本点分布图
基于自然资源部标准地图服务系统审图号GS(2024)0650号标准地图制作,底图无修改。下同
Figure 1. Distribution map of selected sample points of Cupressus gigantea
The map was based on the standard map with review number GS (2024) 0650 downloaded from the standard map service system of the Ministry of Natural Resources, and the base map was not modified. Same below.
![]()
图 2 最终模型的受试者工作曲线(ROC)验证
Figure 2. Receiver operating curve (ROC) validation of final model
![]()
图 3 主导环境变量的响应曲线示意图(A~E)以及影响巨柏潜在分布的各环境变量的折刀测验重要性(F)
Figure 3. Response curve diagram of dominant environmental variables (A–E) and importance of environmental variables affecting potential distribution of Cupressus gigantea based on Jackknife test (F)
![]()
图 4 巨柏在我国的潜在适宜区分布
Figure 4. Distribution of potential suitable habitats for Cupressus gigantea in China
表 1 本研究所用的环境变量
Table 1 Environmental variables used in this study
类型
Type代码
Code描述
Description单位
Unit说明
Expression气候 bio1 年平均温度 ℃ 等温性bio3=(bio2 / bio7)×100,温度的年较差bio7=bio5−bio6,反映温差特点;温度季节性bio4=温度标准差×100,使用温度的标准偏差表示,反映平均温度的变化幅度;降水季节性bio15采用降水的变异系数表示,反映降水量的季节性分布。
温度和降水通常是影响植物生长发育、繁殖扩散的重要因子。巨柏具有喜温、喜光、趋湿、抗旱、抗寒等生态特性[32]。bio2 每月最高温与最低温差值的平均值 ℃ bio3 等温性 % bio4 温度季节性 − bio5 最热月的最高温度 ℃ bio6 最冷月的最低温度 ℃ bio7 温度的年较差 ℃ bio8 最湿季度的平均温度 ℃ bio9 最干季度的平均温度 ℃ bio10 最热季度的平均温度 ℃ bio11 最冷季度的平均温度 ℃ bio12 年降水量 mm bio13 最湿月的降水 mm bio14 最干月的降水量 mm bio15 降水量季节性变异系数 − bio16 最湿季度的降水量 mm bio17 最干季度的降水量 mm bio18 最热季度的降水量 mm bio19 最冷季度的降水量 mm srad 月均太阳辐射 kJ·m−2·d−1 dem 海拔 m 地形 slope 坡度 ° 地形通过对水热条件的调节和再分配影响物种的组成与分布[33]。 aspect 坡向 − 土壤 thickness 土壤厚度 cm 植物的生息与土壤的结构和理化性质紧密相关。相关研究表明土壤质地、养分、密度、含水量等是影响巨柏群落分布的主要因子[16]。 bd 土壤容重 g/cm3 ph 土壤pH值 − fcm 田间持水量 cm3/cm3 soc 土壤有机碳含量 g/kg tn 土壤全氮含量 g/kg clay 土壤黏粒含量 % slit 土壤粉粒含量 % sand 土壤砂粒含量 % 水文 dr 与河流的距离 m 水线地类是巨柏种子萌发、幼苗幼树构建的重要地类[6]。研究认为水或水中动物可能是巨柏沿雅鲁藏布江分布传播的重要媒介[4]。 人类活动 hf 人类足迹指数 − 伐木、樵采、祭祀拜佛等人类活动是影响巨柏分布的重要因子[4]。 植被 NDVI 归一化植被指数 − 巨柏的更新状况会受灌木层郁闭度和竞争程度的影响[6, 34]。  下载: 导出CSV
下载: 导出CSV
表 2 MaxEnt模型“kuenm”包优化评价指标及最优模型参数配置
Table 2 Evaluation metrics of MaxEnt model generated by kuenm and optimal model parameter configuration
模型
Model平均AUC比值
Mean_AUC_ratiopROC统计显著性
pval_pROC10%遗漏率
Omission_rate_at_10%AICc差值
delta_AICc参数数量
num_parametersM_1_F_QT * 1.999 0 0.216 0.000 22 M_1_F_T 1.999 0 0.297 0.933 18 M_1_F_LQPTH 1.999 0 0.189 55.277 38 注:加粗字体表示“kuenm”包筛选的优化模型,未加粗字体表示默认参数模型; * 指本研究确定的最优模型。 Notes: Bold entries represent optimized model parameters selected by “kuenm” package; non-bold entry indicates default parameter model; * signifies optimal model identified in this study.
下载: 导出CSV
表 3 主要环境变量的贡献率及重要性分析
Table 3 Analysis of contribution rate and importance of main environmental variables
环境变量
Environmental variables描述
Description贡献率
Percent contribution / %置换重要性
Permutation importancedr 与河流的距离 19.47 0.07 bio4 温度季节性 18.42 62.16 bio3 等温性 18.23 0.00 slope 坡度 14.11 0.10 bio19 最冷季度的降水量 10.67 0.85 srad 月均太阳辐射 5.68 1.95 dem 海拔 4.02 0.11 bio15 降水季节性 3.14 0.82 fcm 田间持水量 3.05 0.68 ph05 0~5 cm土层的pH值 1.54 2.38 bio11 最冷季度的平均温度 0.94 29.99
下载: 导出CSV
表 4 全国巨柏主要潜在适宜分布区面积统计
Table 4 Statistical details regarding potential suitable distribution areas for C. gigantea in China
行政区等级
Administrative
region位置
Location非适宜区
Unsuitable zone / km2低适宜区
Lowly suitable zone / km2中适宜区
Moderately suitable zone / km2高适宜区
Highly suitable zone / km2省级 西藏自治区 1 127 185.00 20 574.00 2 298.00 746.00 四川省 484 766.00 67.00 − − 市级 林芝市 98 206.00 3 629.00 1 116.00 563.00 山南市 65 159.00 8 675.00 1 120.00 183.00 拉萨市 25 356.00 3 234.00 34.00 − 日喀则市 167 216.00 5 031.00 28.00 − 甘孜藏族自治州 148 495.00 65.00 − − 那曲市 340 058.00 5.00 − − 凉山彝族自治州 60 227.00 2.00 − −
下载: 导出CSV
-
[1] 郑万钧,傅立国,诚静容. 中国裸子植物[J]. 中国科学院大学学报,1975,13(4):56−123. [2] 司琳杰. 西藏珍稀植物巨柏的研究发展概论[J]. 绿色科技,2015(5):45−47. doi: 10.3969/j.issn.1674-9944.2015.05.021 [3] 郑维列,薛会英,罗大庆,王景升,罗建. 巨柏种群的生态地理分布与群落学特征[J]. 林业科学,2007,43(12):8−15. doi: 10.3321/j.issn:1001-7488.2007.12.002 Zheng WL,Xue HY,Luo DQ,Wang JS,Luo J. Eco-geographic distribution and coenology characteristics of Cupressus gigantea[J]. Scientia Silvae Sinicae,2007,43(12):8−15. doi: 10.3321/j.issn:1001-7488.2007.12.002
[4] 尹金迁,赵垦田. 西藏高原巨柏的研究进展与展望[J]. 林业与环境科学,2019,35(2):116−122. Yin JQ,Zhao KT. Research history and prospect of Cupressus gigantean in Tibetan Plateau[J]. Forestry and Environmental Science,2019,35(2):116−122.
[5] 陈端. 西藏巨柏的研究现状与前景[J]. 西藏科技,1995(2):7−11. [6] 李永霞. 珍稀濒危树种巨柏天然更新研究[D]. 拉萨:西藏大学,2023:3−4. [7] Warren DL,Seifert SN. Ecological niche modeling in Maxent:the importance of model complexity and the performance of model selection criteria[J]. Ecol Appl,2011,21(2):335−342. doi: 10.1890/10-1171.1
[8] Phillips SJ,Anderson RP,Schapire RE. Maximum entropy modeling of species geographic distributions[J]. Ecol Model,2006,190(3-4):231−259. doi: 10.1016/j.ecolmodel.2005.03.026
[9] 邢丁亮,郝占庆. 最大熵原理及其在生态学研究中的应用[J]. 生物多样性,2011,19(3):295−302. doi: 10.3724/SP.J.1003.2011.08318 Xing DL,Hao ZQ. The principle of maximum entropy and its applications in ecology[J]. Biodiversity Science,2011,19(3):295−302. doi: 10.3724/SP.J.1003.2011.08318
[10] Elith J,Graham CH,Anderson RP,Dudík M,Ferrier S,et al. Novel methods improve prediction of species’ distributions from occurrence data[J]. Ecography,2006,29(2):129−151. doi: 10.1111/j.2006.0906-7590.04596.x
[11] 蔡静芸,张明明,粟海军,张海波. 生态位模型在物种生境选择中的应用研究[J]. 经济动物学报,2014,18(1):47−52,58. Cai JY,Zhang MM,Su HJ,Zhang HB. Application of ecological niche models for selection of species habitat[J]. Journal of Economic Animal,2014,18(1):47−52,58.
[12] Mahmoodi S,Heydari M,Ahmadi K,Khwarahm NR,Karami O,et al. The current and future potential geographical distribution of Nepeta crispa Willd.,an endemic,rare and threatened aromatic plant of Iran:implications for ecological conservation and restoration[J]. Ecol Indic,2022,137:108752.
[13] Yang JT,Huang Y,Jiang X,Chen H,Liu M,Wang RL. Potential geographical distribution of the edangred plant Isoetes under human activities using MaxEnt and GARP[J]. Glob Ecol Conserv,2022,38:e02186.
[14] Yang H,Li JL,Milne RI,Tao WJ,Wang Y,et al. Genomic insights into the genotype–environment mismatch and conservation units of a Qinghai–Tibet Plateau endemic cypress under climate change[J]. Evol Appl,2022,15(6):919−933. doi: 10.1111/eva.13377
[15] Lei P,Liu Z,Li JX,Jin GZ,Xu LP,et al. Integration of the physiology,transcriptome and proteome reveals the molecular mechanism of drought tolerance in Cupressus gigantea[J]. Forests,2022,13(3):401. doi: 10.3390/f13030401
[16] 杨刚刚,郝海霞,岳艳鹏,周继华,来利明,郑元润. 雅鲁藏布江巨柏分布区植物群落特点、分布与环境解释[J]. 北京师范大学学报(自然科学版),2023,59(1):156−162. doi: 10.12202/j.0476-0301.2022125 Yang GG,Hao HX,Yue YP,Zhou JH,Lai LM,Zheng YR. Community characteristics,distribution and environment interpretation of Cupressus gigantea along Yarlung Zangbo River in Tibet[J]. Journal of Beijing Normal University (Natural Science),2023,59(1):156−162. doi: 10.12202/j.0476-0301.2022125
[17] 姚祺. 巨柏移植前生长调控的研究[D]. 林芝:西藏大学农牧学院,2021:32−57. [18] 常馨月. 巨柏生殖特征及发育机制研究[D]. 拉萨:西藏大学,2023:75−131. [19] 姚祺,李佶芸,赵垦田. 基于MaxEnt模型的巨柏青藏高原生态适宜性研究[J]. 高原农业,2021,5(2):109−114. Yao Q,Li JY,Zhao KT. Research on ecological suitability of giant cypress on Qinghai-Tibet Plateau based on MaxEnt model[J]. Journal of Plateau Agriculture,2021,5(2):109−114.
[20] Brown JL. SDMtoolbox:a python-based GIS toolkit for landscape genetic,biogeographic and species distribution model analyses[J]. Methods Ecol Evol,2014,5(7):694−700. doi: 10.1111/2041-210X.12200
[21] 王运生. 生态位模型在外来入侵物种风险评估中的应用研究[D]. 长沙:湖南农业大学,2007:4,26−27. [22] Li XP,Liu X,Kraus F,Tingley R,Li YM. Risk of biological invasions is concentrated in biodiversity hotspots[J]. Front Ecol Environ,2016,14(8):411−417. doi: 10.1002/fee.1321
[23] 曹倩,高庆波,郭万军,张毓,王智华,等. 基于MaxEnt模拟人类活动与环境因子对青藏高原特有植物祁连獐牙菜潜在分布的影响[J]. 植物科学学报,2021,39(1):22−31. Cao Q,Gao QB,Guo WJ,Zhang Y,Wang ZH,et al. Impacts of human activities and environmental factors on potential distribution of Swertia przewalskii Pissjauk. ,an endemic plant in Qing-Tibetan Plateau,using MaxEnt[J]. Plant Science Journal,2021,39(1):22−31.
[24] Lehner B,Grill G. Global river hydrography and network routing:baseline data and new approaches to study the world's large river systems[J]. Hydrol Process,2013,27(15):2171−2186. doi: 10.1002/hyp.9740
[25] Bowen AKM,Stevens MHH. Temperature,topography,soil characteristics,and NDVI drive habitat preferences of a shade-tolerant invasive grass[J]. Ecol Evol,2020,10(19):10785−10797. doi: 10.1002/ece3.6735
[26] Liu F,Wu HY,Zhao YG,Li DC,Yang JL,et al. Mapping high resolution National Soil Information Grids of China[J]. Sci Bull,2022,67(3):328−340. doi: 10.1016/j.scib.2021.10.013
[27] 边喜丽,杨小林,马和平,赵垦田. 藏东南巨柏根系结构特征分析[J]. 西部林业科学,2017,46(4):107−112. Bian XL,Yang XL,Ma HP,Zhao KT. Root structure and specific characteristics of Cupressus gigantea in Southeast Tibet[J]. Journal of West China Forestry Science,2017,46(4):107−112.
[28] Venter O,Sanderson EW,Magrach A,Allan JR,Beher J, et al. Last of the wild project,version 3 (LWP-3):2009 human footprint,2018 release (version 2018.00)[R]. Palisades:NASA Socioeconomic Data and Applications Center (SEDAC),2018.
[29] De Marco Júnior P,Nóbrega CC. Evaluating collinearity effects on species distribution models:an approach based on virtual species simulation[J]. PLoS One,2018,13(9):e0202403. doi: 10.1371/journal.pone.0202403
[30] Raghavan RK,Barker SC,Cobos ME,Barker D,Teo EJM,et al. Potential spatial distribution of the newly introduced long-horned tick,Haemaphysalis longicornis in North America[J]. Sci Rep,2019,9(1):498. doi: 10.1038/s41598-018-37205-2
[31] Jayasinghe SL,Kumar L. Modeling the climate suitability of tea [Camellia sinensis(L.) O. Kuntze] in Sri Lanka in response to current and future climate change scenarios[J]. Agric For Meteorol,2019,272-273:102−117. doi: 10.1016/j.agrformet.2019.03.025
[32] 李乾振,李朝晖. 闽东引种巨柏育苗试验初报[J]. 林业科学,2003,39(S1):184−186. Li QZ,Li ZH. Preliminary report on seedling raising experiment for introducing Cupressus gigantea in eastern Fujian Province[J]. Scientia Silvae Sinicae,2003,39(S1):184−186.
[33] 郑诚,温仲明,郭倩,樊勇明,杨玉婷,高飞. 基于MaxEnt模型的延河流域草本植物适生分布与功能性状分析[J]. 生态学报,2021,41(17):6825−6835. Zheng C,Wen ZM,Guo Q,Fan YM,Yang YT,Gao F. Analysis of suitability distribution and functional traits of common herb species in Yanhe River catchment based on MaxEnt model[J]. Acta Ecologica Sinica,2021,41(17):6825−6835.
[34] 吴兴,赵南先,段代祥,陈贻竹. 西藏特有珍稀植物巨柏的研究进展与展望[J]. 福建林业科技,2005,32(3):160−164. doi: 10.3969/j.issn.1002-7351.2005.03.043 Wu X,Zhao NX,Duan DX,Chen YZ. Research progress and expectation of Cupressus gigantean,the rare and endemic plant in Xizang[J]. Journal of Fujian Forestry Science and Technology,2005,32(3):160−164. doi: 10.3969/j.issn.1002-7351.2005.03.043
[35] Cobos ME,Peterson AT,Barve N,Osorio-Olvera L. kuenm:an R package for detailed development of ecological niche models using Maxent[J]. PeerJ,2019,7:e6281. doi: 10.7717/peerj.6281
[36] Phillips SJ,Dudík M. Modeling of species distributions with Maxent:new extensions and a comprehensive evaluation[J]. Ecography,2008,31(2):161−175. doi: 10.1111/j.0906-7590.2008.5203.x
[37] Chen BR,Zou H,Zhang BY,Zhang XY,Jin XX,et al. Distribution pattern and change prediction of Saposhnikovia divaricata suitable area in China under climate change[J]. Ecol Indic,2022,143:109311. doi: 10.1016/j.ecolind.2022.109311
[38] Swets JA. Measuring the accuracy of diagnostic systems[J]. Science,1988,240(4857):1285−1293. doi: 10.1126/science.3287615
[39] 王运生,谢丙炎,万方浩,肖启明,戴良英. ROC曲线分析在评价入侵物种分布模型中的应用[J]. 生物多样性,2007,15(4):365−372. doi: 10.3321/j.issn:1005-0094.2007.04.005 Wang YS,Xie BY,Wan FH,Xiao QM,Dai LY. Application of ROC curve analysis in evaluating the performance of alien species’ potential distribution models[J]. Biodiversity Science,2007,15(4):365−372. doi: 10.3321/j.issn:1005-0094.2007.04.005
[40] Liu CR,Berry PM,Dawson TP,Pearson RG. Selecting thresholds of occurrence in the prediction of species distributions[J]. Ecography,2005,28(3):385−393. doi: 10.1111/j.0906-7590.2005.03957.x
[41] The Core Writing Team,Pachauri RK,Reisinger A. Climate Change 2007:Synthesis Report. Contribution of Working Groups Ⅰ,Ⅱ and Ⅲ to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change[M]. Geneva:IPCC,2007:27,83.
[42] Wang RL,Li Q,He SS,Liu Y,Wang MT,Jiang G. Modeling and mapping the current and future distribution of Pseudomonas syringae pv. actinidiae under climate change in China[J]. PLoS One,2018,13(2):e0192153. doi: 10.1371/journal.pone.0192153
[43] Tang CQ,Matsui T,Ohashi H,Dong YF,Momohara A,et al. Identifying long-term stable refugia for relict plant species in East Asia[J]. Nat Commun,2018,9(1):4488. doi: 10.1038/s41467-018-06837-3
[44] 扎西次仁. 西藏巨柏(Cupressus gigantea)的遗传多样性与精油化学成分变异及其保护生物学意义[D]. 上海:复旦大学,2008:71. [45] 王景升,郑维列,潘刚. 巨柏种子活力与濒危的关系[J]. 林业科学,2005,41(4):37−41. doi: 10.3321/j.issn:1001-7488.2005.04.007 Wang JS,Zheng WL,Pan G. Relation between being endangered and seed vigor about Cupressus gigantea in Tibet[J]. Scientia Silvae Sinicae,2005,41(4):37−41. doi: 10.3321/j.issn:1001-7488.2005.04.007
[46] 徐海燕. 中国柏木属(Cupressus L.)四个物种的遗传多样性研究[D]. 兰州:兰州大学,2011:28. -
其他相关附件
-
PDF格式
2. 曾伟英 附表1和附图1 点击下载(247KB)
-
计量
- 文章访问数: 32
- HTML全文浏览量: 6
- PDF下载量: 7
