
Phenotypic characteristics and comprehensive evaluation of high-quality germplasm resources of Gentiana rigescens Franch. ex Hemsl.
-
摘要:
为探讨野生滇龙胆(Gentiana rigescens Franch. ex Hemsl.)优质高产植株性状特征,以采自不同地区的877株滇龙胆为研究材料,利用主成分分析(PCA)、层次聚类(HCA)、隶属函数等方法对有效成分产量性状进行评价。结果显示,17个性状中,根部龙胆苦苷产量多样性指数最高,叶部马钱苷酸和当药苷产量的多样性指数较低;结合隶属函数对所有样品进行评分,发现优质高产种源共214株,占总样品数的24.40%,分布于云南、四川和贵州;变量投影重要性准则(VIP)分析表明,云南与四川的优质种源主要性状特征较为接近,均为当药苷、马钱苷酸及6'-O-β-D-葡萄糖基龙胆苦苷高产;贵州优质种源则为獐牙菜苦苷高产。基于3种机器学习算法建立不同等级种源的鉴别模型,结果发现随机森林(RF)算法建立的判别模型的预测精度和稳定性较高,能对不同等级种源进行有效划分。
Abstract:We explored the phenotypic characteristics and established a classification strategy of high-quality germplasm resources of wild Gentiana rigescens Franch. ex Hemsl. In total, 887 samples of G. rigescens collected from different regions were used as research materials. Principal component analysis (PCA), hierarchical clustering analysis (HCA), and membership function analysis were used to analyze and evaluate 17 active ingredient yield traits of the roots, stems, and leaves. Results showed that gentiopicroside yield in the roots had the highest Shannon-Wiener index value (I = 1.64), while loganic acid and sweroside acid yields in the leaves had the lowest I values (I = 0.73). Based on D value scoring and membership function analysis, we identified 214 high-quality and high-yield seed sources, accounting for 24.40% of the total sample size, distributed in Yunnan, Sichuan, and Guizhou. Variable importance in projection (VIP) analysis showed similar phenotypic characteristics among the high-quality germplasms in Yunnan and Sichuan, which were characterized by high sweroside, loganic acid, and 6'-O-β-D-glucopyranosylgentiopicroside yield. The high-quality germplasms in Guizhou were characterized by high swertiamarin yield. Among the three different machine learning algorithms, results showed that the discrimination model established using the Random Forest (RF) algorithm had the highest prediction accuracy and stability and could effectively identify different provenances.
-
优良种质资源是保障中药材生产、实现药材品质提升与种质创新的前提和基础[1-3]。中药历史悠久,但其药物开发利用多以野生资源为主,药材驯化栽培水平远落后于传统农作物[3-5]。现代社会对中药及相关产品的市场需求增加,推动了中药材规模化种植业的发展[3, 4]。在此过程中,科学筛选优良种源,建立全面、客观的种源筛选与评价策略,对推动优质中药材生产与生态种植具有重要意义[4, 5]。
滇龙胆(Gentiana rigescens Franch. ex Hemsl.)在西南地区入药历史悠久,是传统保肝中药龙胆的主要基原植物,也是近年推动“云药”产业发展的关键物种[6-8]。滇龙胆作为“十大云药”重要成员(http://www.satcm.gov.cn/),以其为原料的中成药已有百余种,市场对其需求量逐年增加,野生资源难以提供稳定供给[6]。因此,药材规模化种植成为滇龙胆可持续利用的关键,其优良种源的筛选也迫在眉睫。
当前滇龙胆种源筛选、质量评价指标(有效成分含量)和产量评价指标(药用部位干重)相对独立,缺乏适宜的综合评价方法[9-11]。此外,植物化学与药理学研究显示,除《中国药典》规定的龙胆苦苷(Gentiopicroside,Gentio)外,6'-O-β-D-葡萄糖基龙胆苦苷(6'-O-β-D-glucopyranosylgentiopicroside,6'-O-β-D-gluco)、马钱苷酸(Loganic acid,LA)、獐牙菜苦苷(Swertiamarin,Swer)、当药苷(Sweroside,Swero)、异荭草素(Isoorientin,Isoo)等化学成分均是滇龙胆发挥药效的物质基础[7, 12],这些化学成分也应作为药材种源质量的评价指标[13]。作为一种主要生长在山区的药材,滇龙胆分布区土地稀缺,药材种植面临与粮食生产用地冲突的困境[14, 15]。从提升药材整体药用价值、提高土地利用效率、增加单株产量角度出发,筛选有效成分含量高的优质高产种源,不仅能保证滇龙胆药材品质,提高药材生产效率,也有助于节约山区土地资源,契合中药产业循环利用与绿色生产的发展理念[3, 16, 17]。
本研究基于对云南、四川、贵州等地野生滇龙胆种质资源的系统调查,采集了877株个体根、茎、叶有效成分产量性状的数据,结合主成分分析(Principal component analysis,PCA)、隶属函数(Membership function)等方法,综合分析不同地理来源植株的性状指标,确定高产优质种源的性状特征,开展滇龙胆种质资源质量等级评价。同时运用偏最小二乘判别分析(Partial least squares-discriminant analysis,PLS-DA)、正交偏最小二乘判别(Orthogonal partial least squares-discriminant analysis,OPLS-DA)和随机森林(Random forest,RF)等多种机器学习算法建立不同等级种源的分类鉴别模型。研究结果旨在为野生滇龙胆优良种源筛选及分级策略的制定提供理论依据。
1. 材料与方法
1.1 实验材料
研究基于课题组2016-2019年对云南、四川、贵州、广西、湖南等5省22个州市的资源调查,共获取877株野生滇龙胆单株用于本文分析评价(样品信息见附表1
1 )。种源地为云南的滇龙胆共720株,其中,采自滇西北、滇西、滇西南、滇南、滇中、滇东和滇东南的样本分别为63、138、30、56、350、49和34株。种源地为四川的滇龙胆共49株,均采自川西南。种源地为贵州的滇龙胆共88株,其中30株采自黔西南,28株采自黔南,20株采自黔中,10株采自黔东南。其余20株样品分别来源于广西(10株)和湖南(10株)。样品采集时间为每年11月药材传统采收期。每个采集地间隔大于30 km,同一地点采集植株个体数尽量大于10株,单株间距 ≥ 10 m。样品采集均挑选开花植株;采集后详细记录采集地信息,用去离子水快速洗净植株,带回实验室,放置于50 ℃条件下烘干至恒重,用电子天平称量根、茎、叶部位的干重;随后粉碎,过60目筛,用于有效成分含量检测,并进一步结合各部位干重计算根、茎、叶的有效成分产量。
1.2 产量性状测量
通过测量和计算共获取17个产量性状(附表2
2 、附表32 )。每株滇龙胆不同部位马钱苷酸、龙胆苦苷、獐牙菜苦苷、当药苷、6'-O-β-D-葡萄糖基龙胆苦苷以及异荭草素等的产量计算方法如下:Y=C×B (1) 式中,Y为某一有效成分产量,C为有效成分含量,B为部位干重(根、茎或叶)。6种有效成分含量依据课题组开发方法测定(有效成分色谱图详见附图1
2 )[10]。1.3 性状统计与综合评分计算
用Shannon多样性指数评估滇龙胆样品根、茎、叶产量性状的多样性[18]。利用主成分分析探索有效成分产量数据的分离与聚散趋势[19];使用层次聚类对样品进行分类研究[20]。
依据公式(2)计算各性状隶属函数值;根据公式(3)结合主成分分析结果计算各性状权重;最后将所有性状的隶属函数值与性状权重代入公式(4)中,计算每株样品产量性状的综合评分[21, 22]:
M(Xi)=(Xi−Ximin)/(Ximax−Ximin) (2) 式中,M(Xi)为样品第i个性状的隶属函数值,Xi为每株个体第i个性状的实测值,Xi min和Xi max分别表示所有样品第i个性状的最小值和最大值。
Wi=Pi/n∑i=1Pi (3) 式中Wi为样品第i个性状的权重,Pi是经过主成分分析计算得到的第i个性状的贡献率。
D=n∑i=1[M(Xi)×Wi] (4) 式中,D值为每株个体11个性状的综合得分,i表示参与评分的第i个性状。
偏最小二乘判别分析结合变量投影重要性准则(Variable important in projection,VIP)分析,用于计算产量性状的VIP值,筛选出导致不同分布区优质种源分类差异的关键性状(VIP值大于1.00)[23]。
1.4 建模分析
依据样品种源分级结果,将产量性状设为建模变量,利用偏最小二乘判别、正交偏最小二乘判别和随机森林3种算法建立不同等级样品的判别模型[13, 24]。
数据集70%作为训练集,30%作为测试集,使用正确率、灵敏度、特异性和马修斯相关系数对3种算法建立的模型进行评价。利用R2X、Q2、校正均方根误差、交叉验证均方根误差和置换重要性检验进一步评价PLS-DA和OPLS-DA模型[13, 24]。
1.5 统计软件
性状数值统计用SPSS 25.0 软件计算,层次聚类(Hierarchical clustering,HCA)、VIP、PCA分析及PLS-DA、OPLS-DA建模用SIMCA 14.1软件运算;RF算法建模由R包randomForest、varSelRF和ParallelForest共同完成;使用Origin 20.0软件绘图。
2. 结果与分析
2.1 滇龙胆产量性状的多样性分析
877株滇龙胆根、茎、叶有效成分产量性状(附表2)的Shannon多样性指数(I)计算结果显示(表1),各性状多样性指数在0.73~1.64,平均值为1.20。所有性状中,根部龙胆苦苷产量的多样性指数(I = 1.64)最高;根部獐牙菜苦苷产量(I = 1.53)和茎部当药苷产量(I = 1.56)次之;叶部马钱苷酸产量和当药苷产量多样性指数最低,均为0.73。
表 1 滇龙胆有效成分产量性状的多样性指数Table 1. Shannon diversity index of yield traits of active ingredients in Gentiana rigescens产量性状
Yield traitShannon多样性指数(I) Shannon diversity index 根Root 茎Stem 叶Leaf 马钱苷酸 1.51 0.94 0.73 6'-O-β-D-葡萄糖基龙胆苦苷 1.39 1.15 1.02 獐牙菜苦苷 1.53 1.20 1.09 龙胆苦苷 1.64 1.35 1.05 当药苷 1.37 1.56 0.73 异荭草素 − 1.08 1.05 各类有效成分中,龙胆苦苷产量多样性指数均值最高(根、茎、叶性状多样性指数均值为1.35),马钱苷酸和异荭草素产量多样性指数均值最低(两种成分根、茎和叶性状多样性指数均值均为1.06)。
各部位中,根部性状多样性指数均值最高(1.49),茎部次之(1.21),叶部有效成分产量性状的多样性指数均值最低(0.94)。
2.2 基于药材产量性状的主成分分析
利用无监督的主成分分析算法结合根、茎、叶有效成分产量性状(共17个),研究877株滇龙胆的聚散趋势。分析结果显示,第1主成分贡献率为44.816%,第2主成分贡献率为15.885%,第3主成分贡献率为9.423%,第4和第5主成分贡献率分别为6.982%和4.592%。前5个主成分累计贡献率大于80.000%,表明这5个主成分可代表17个性状的大部分信息(表2)。
表 2 基于滇龙胆17个产量性状的主成分分析Table 2. Principal component analysis results based on 17 yield traits of Gentiana rigescens成分
Component初始特征值Eigenvalue 特征值
Eigenvalue方差贡献率
Contribution of variance / %累积方差贡献率
Cumulative contribution / %1 7.619 44.816 44.816 2 2.700 15.885 60.700 3 1.602 9.423 70.123 4 1.187 6.982 77.105 5 0.781 4.592 81.697 依据成分矩阵的特征向量(附表3
3 )可知,第1主成分代表根部有效成分产量变化,其所属向量指标分别为:根部马钱苷酸产量、根部6'-O-β-D-葡萄糖基龙胆苦苷产量、根部獐牙菜苦苷产量、根部龙胆苦苷产量和根部当药苷产量。第2主成分代表茎部有效成分的产量变化,其所属向量指标分别为:茎部马钱苷酸产量、茎部6'-O-β-D-葡萄糖基龙胆苦苷产量、茎部獐牙菜苦苷产量、茎部龙胆苦苷产量、茎部当药苷产量。第3主成分主要代表叶部产量性状,其所属向量指标包括:叶部马钱苷酸产量、叶部6'-O-β-D-葡萄糖基龙胆苦苷产量、叶部獐牙菜苦苷产量、叶部龙胆苦苷产量和叶部异荭草素产量。而第4和第5主成分所属向量分别为叶部当药苷产量和茎部异荭草素产量。依据主成分得分,所有样品可聚为4类(图1)。采自云南、四川和贵州的滇龙胆在每类中均有分布;采自广西的滇龙胆主要分布在第Ⅱ类,采自湖南的植株主要分布于第Ⅱ类和第Ⅳ类。
2.3 性状权重分析与综合评价
依据性状数值结合主成分分析、层次聚类发现,样品呈现一定的聚类趋势,但难以对各类样品的特征进行科学描述,因此需应用更加综合的评价策略。在滇龙胆17个性状主成分分析基础上,计算所有性状的隶属函数值与主成分权重系数(主成分1~5的权重系数分别为:0.549、0.194、0.115、0.085和0.056),最终获得每株滇龙胆的综合得分D值。
877株滇龙胆综合得分统计结果显示(附表4
3 ),样品D值范围在0.01~2.34,大部分样品综合得分集中分布于0.13~0.44。评分排名前20的样品主要来源于云南昆明、大理、曲靖、迪庆、贵州黔南等地;综合得分排序后20名的样品中,云南文山和广西百色地区的样品合计占了35%。此外,云南楚雄、丽江、红河、玉溪及四川凉山、攀枝花,贵州黔西南等地也有得分较低的样品(附表53 )。依据样品D值的上四分位数和下四分位数(附表2),将所有样品划分为3个等级。等级1:D值 > 0.44,代表有效成分产量高的优质种源;等级2:0.13 ≤ D ≤ 0.44值,代表普通种源;等级3:D < 0.13,代表有效成分产量较低的劣质种源。
2.4 优质种源的区域性状特征
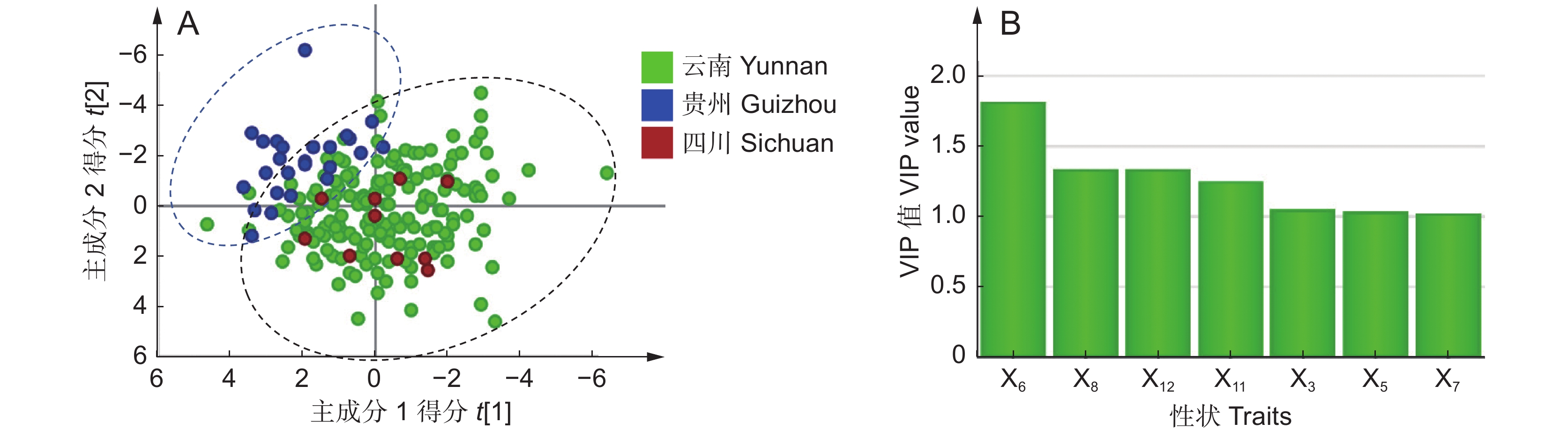
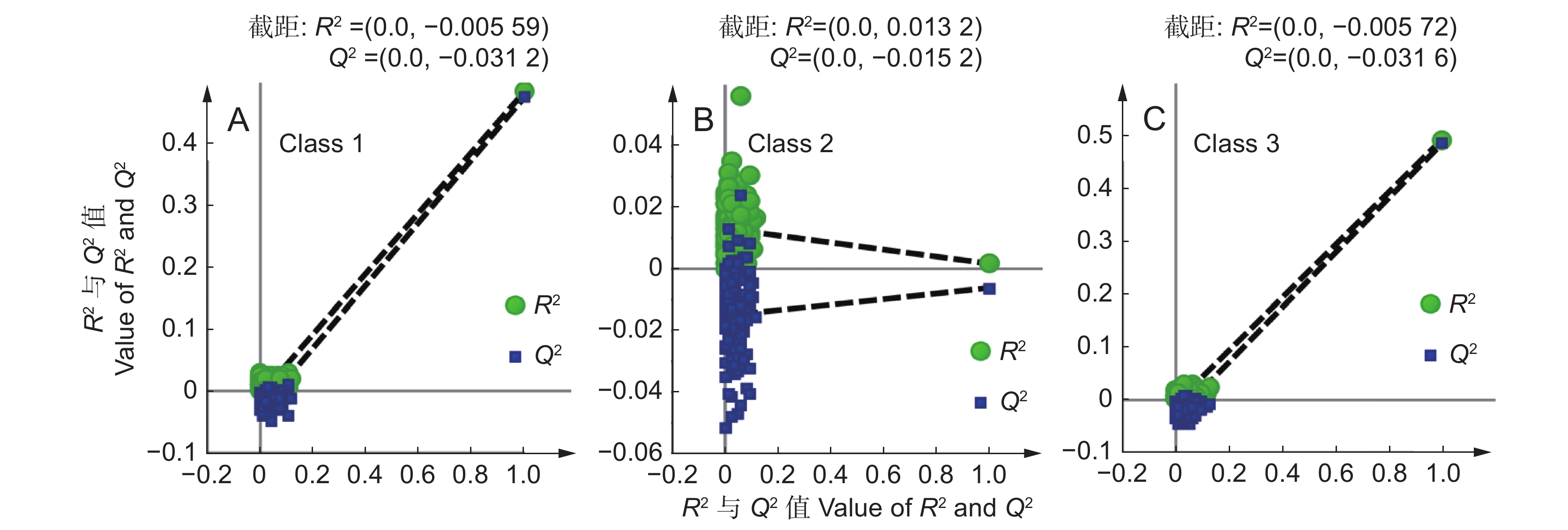
为分析云南、四川、贵州等地滇龙胆优质种源的性状特征,利用有监督的偏最小二乘-判别分析结合17个产量性状(X1~X17),对上述地区综合评分D值 ˃ 0.44的样品进行分类。得分图显示(R2X = 0.616,Q2 = 0.263),所有样品总体聚为两类(图2:A);其中云南和四川的滇龙胆性状最接近,样品难以区分;采自贵州与其他地区的滇龙胆样品之间有较明显的性状差异。
![]() 图 2 滇龙胆样品偏最小二乘-判别分析得分图(A)与VIP分析图(B)Figure 2. Loading plot based on PLS-DA (A) and VIP analysis (B) of Gentiana rigescens samples
图 2 滇龙胆样品偏最小二乘-判别分析得分图(A)与VIP分析图(B)Figure 2. Loading plot based on PLS-DA (A) and VIP analysis (B) of Gentiana rigescens samplesVIP分析结果显示(图2:B),导致两类样品差异的主要性状指标依次为:茎部马钱苷酸产量(X6:VIP = 1.82)、茎部獐牙菜苦苷产量(X8:VIP = 1.34)、叶部位马钱苷酸产量(X12:VIP = 1.34)、茎部异荭草苷产量(X11:VIP = 1.25)、根部獐牙菜苦苷产量(X3:VIP = 1.05)、根部当药苷产量(X5:VIP = 1.04)及茎部6'-O-β-D-葡萄糖基龙胆苦苷产量(X7:VIP = 1.02)。进一步比较性状均值(附表6
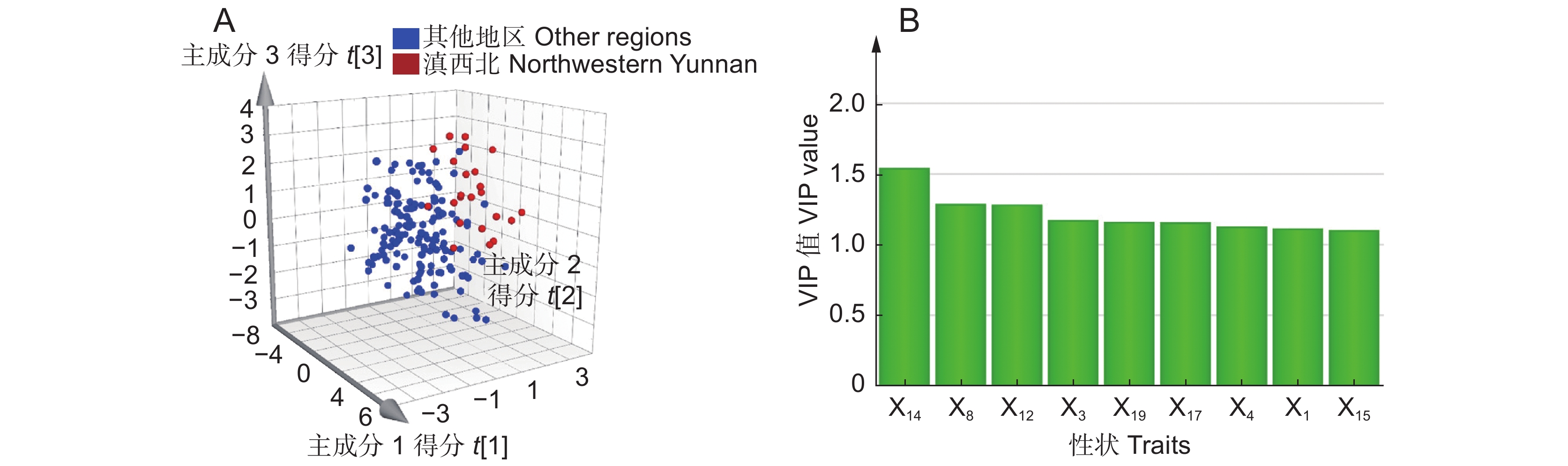
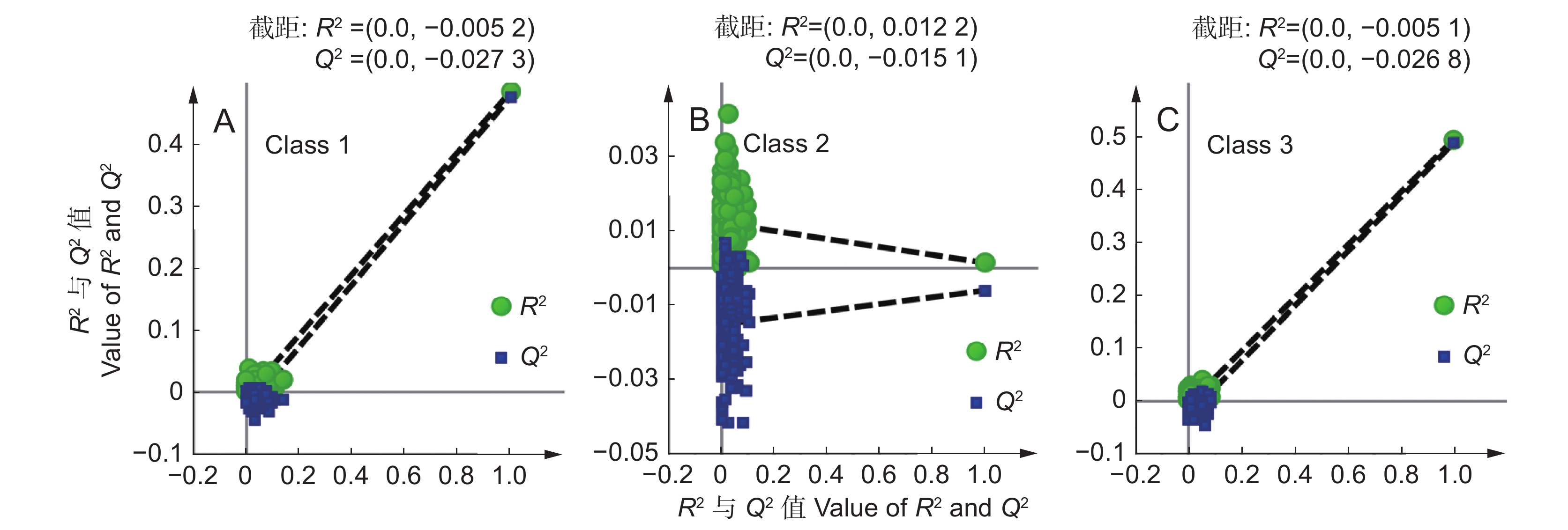
4 ),结果显示,云南、四川滇龙胆优质种源的根部当药苷、茎部和叶部马钱苷酸、茎部6'-O-β-D-葡萄糖基龙胆苦苷产量较高;贵州滇龙胆优质种源的根、茎部獐牙菜苦苷与茎部异荭草素产量高。云南是滇龙胆优质种源数量最多的省份,对云南境内优质种源性状特征进一步分析发现,不同分布区滇龙胆性状有细微差异(PLS-DA:R2X = 0.644,Q2 = 0.360)。样品大致可分为两类,第1类采自滇西北,第2类采自云南其他地区(图3:A)。VIP分析结果显示(图3:B),导致两组个体差异的重要性状依次为:叶部獐牙菜苦苷产量(X14:VIP = 1.54)、茎部獐牙菜苦苷产量(X8:VIP = 1.27)、叶部马钱苷酸产量(X12:VIP = 1.28)、根部獐牙菜苦苷产量(X3:VIP = 1.17)、叶部异荭草素产量(X17:VIP = 1.16)、根部龙胆苦苷产量(X4:VIP = 1.13)、根部马钱苷酸产量(X1:VIP = 1.11)以及叶部龙胆苦苷产量(X15:VIP = 1.10)。
![]() 图 3 样品偏最小二乘-判别分析得分图(A)与VIP分析图(B)Figure 3. Loading plot of PLS-DA (A) and VIP analysis (B)
图 3 样品偏最小二乘-判别分析得分图(A)与VIP分析图(B)Figure 3. Loading plot of PLS-DA (A) and VIP analysis (B)上述性状比较显示(附表7
5 ),滇西北样品根部的马钱苷酸、獐牙菜苦苷及茎部的獐牙菜苦苷产量略低于云南其他地区,但该地区植株根部龙胆苦苷、叶部马钱苷酸和龙胆苦苷的均值与中位数均明显高于其他地区。2.5 滇龙胆种源分类预测模型的建立
基于877株滇龙胆种质资源的综合评分结果,结合多种机器学习算法,建立滇龙胆种源分类模型,以期为药材优良种源高效筛选与准确鉴别提供方法。建模前,首先将所有样品划分为训练集和测试集。随后将17个根、茎、叶有效成分产量性状设为X变量;利用偏最小二乘、正交偏最小二乘、随机森林3种算法建立不同等级种源的分类模型。
基于偏最小二乘算法建立的分类模型结果显示(表3和附表8
5 ),模型R2X = 0.644,Q2 = 0.360;模型训练集分类正确率在82.24%~92.00%,测试集分类正确率在86.11%~94.05%;建模参数表明模型对变量X的解释能力为64.4%,模型预测结果有待完善和提升。表 3 基于PLS-DA算法的分类建模评价结果Table 3. Evaluation results of classification model based on PLS-DA algorithm评价指标
Evaluation parameter一级
Class 1 / %二级
Class 2 / %三级
Class 3 / %合计
Total / %训练集正确率 92.00 82.24 89.92 88.05 测试集正确率 92.06 86.11 94.05 90.74 注:一级为优质种源,二级为普通种源,三级为劣质种源。下同。 Notes: Classes 1, 2, and 3 refer to high-quality, moderate-quality, and low-quality germplasms, respectively. Same below. 基于正交偏最小二乘算法建立的分类模型参数显示(表4和附表9
5 ),模型R2X = 0.537,Q2 = 0.320;各级样品训练集分类正确率依次为:92.48%、82.24%和89.44%;测试集分类正确率依次为92.86%、86.51%和93.65%。OPLS-DA模型分类预测结果优于PLS-DA模型。表 4 基于OPLS-DA算法的滇龙胆分类模型Table 4. Evaluation results of classification model based on OPLS-DA algorithm评价指标
Evaluation parameter一级
Class 1 / %二级
Class 2 / %三级
Class 3 / %合计
Total / %训练集正确率 92.48 82.24 89.44 88.05 测试集正确率 92.86 86.51 93.65 91.01 建模结果结合200次置换重要性检验结果显示,PLS-DA与OPLS-DA算法建立的分类模型对第二级种源鉴别效果不佳、分类正确率低,且模型预测结果有较明显的过拟合风险(图4:B和图5:B)。
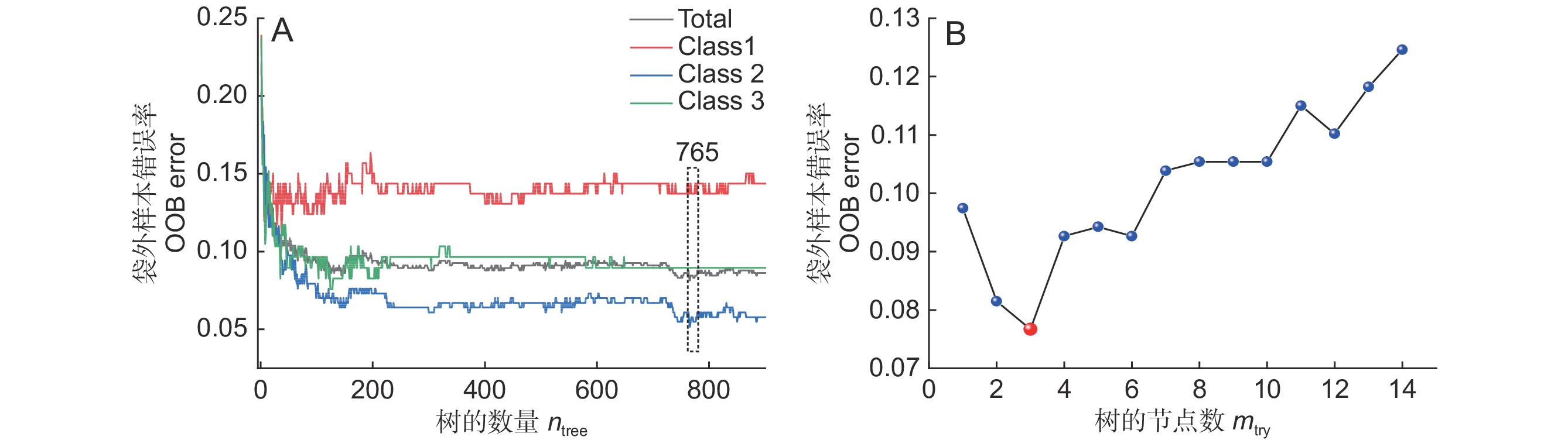
为获得更好的分类模型,尝试运用随机森林算法进行建模研究。模型建立前需确定树的数量(ntree)与树的节点数(mtry)两个建模参数最佳值。通过多次建模计算发现,当ntree= 765(图6:A),mtry= 3时(图6:B),模型OOB-袋外错误率(Out of bag error)最低;因此,利用以上参数建立最终分类模型。
![]() 图 6 随机森林模型ntree(A)与mtry(B)的参数筛选Figure 6. ntree (A) and mtry (B) screening of RF model
图 6 随机森林模型ntree(A)与mtry(B)的参数筛选Figure 6. ntree (A) and mtry (B) screening of RF model建模结果显示(表5和附表10
6 ),RF模型灵敏度、特异性及马修斯相关系数在0.88~0.96。第一级与第三级种源分类正确率均大于95.00%,第二级种源的分类正确率也达到了91.00%以上。模型训练集与测试集样品的总体分类正确率分别为94.14%和94.71%。以上参数均优于PLS-DA和OPLS-DA模型。综合分析认为,3种算法建立的模型中,随机森林算法建模无过拟合风险,各类样品均有较高的分类正确率,属最佳建模算法。表 5 基于随机森林算法建立的滇龙胆分类模型Table 5. Evaluation results of classification model based on RF algorithm评价指标
Evaluation parameter一级
Class 1 / %二级
Class 2 / %三级
Class 3 / %合计
Total / %训练集正确率 95.37 91.21 95.85 94.14 测试集正确率 96.43 92.06 95.63 94.71 3. 讨论
3.1 滇龙胆高产优质种源分级策略的建立
本研究发现,滇龙胆根、茎、叶有效成分产量性状多样性水平较高;根部龙胆苦苷、马钱苷酸、獐牙菜苦苷产量及茎部当药苷产量均呈现丰富的种内变化。这一方面反映出滇龙胆药材产量与质量的不稳定,另一方面也为药材性状改良和人工育种提供了较大空间。
当前滇龙胆质量等级评价主要以根部龙胆苦苷含量为指标[7, 11]。杨天梅等[11]进一步将根长、根粗、根鲜重、干重等产量性状引入到滇龙胆药材质量的分级研究,对《中国药典》评价方法进行了完善。然而上述评价策略主要适用于药材饮片;对药材生产源头—种子、种苗或优良品种仍缺乏全面客观的评价方法。
基于植株表型性状和隶属函数综合评价种源已被广泛应用于小麦(Triticum aestivum L.)、水稻(Oryza sativa L.)等农作物[25-27]。该方法能在兼顾性状多样性的前提下,有效地将复杂性状数据进行简化,更有助于种质资源的客观评价[25-27]。本研究利用有效成分产量这一综合性状,结合主成分分析、隶属函数等方法,对不同地理来源的877株滇龙胆进行综合评分,通过样品D值计算与数值统计,建立了滇龙胆优质高产种源的分级策略。其中一级种源整体特征为全草有效成分产量高;二级种源有效成分产量居中;三级种源全草有效成分产量较低,质量较差。
本研究还发现,不同分布区优质种源具有明显的地域特征。云南、四川的优质高产滇龙胆性状最接近,地上部位有效成分产量与贵州的滇龙胆有较大差异。云南与四川优质种源的主要性状特征为当药苷、马钱苷酸及6'-O-β-D-葡萄糖基龙胆苦苷高产;而贵州优质种源的性状特征为獐牙菜苦苷与异荭草素高产。此外,云南境内高产优质滇龙胆还可进一步划分为滇西北种源与云南其他地区种源;前者最主要的特征为根、叶部位均有较高的有效成分产量。以上结果表明,滇龙胆野生种质资源优良性状呈现丰富的组合与变化,今后还需结合产地环境、种源性状特征、实际生产需求,进行分类利用、定向培育,使滇龙胆优质种源发挥最大的生产效益[5]。
3.2 利用机器学习协助滇龙胆种源的有效分级
机器学习已被广泛应用于植物表型分析与优良性状筛选[28-30]。目前在对滇龙胆资源的评价研究中,PLS-DA、OPLS-DA、RF等算法主要与药材色谱、光谱指纹图谱数据相结合,以解决药材产地溯源问题[31-33]。将机器学习算法应用于种质资源的分级评价尚未见报道。国外学者已利用PLS-DA和人工神经网络(Artificial neural network)对黄龙胆(G. lutea L.)根部样品进行过分级研究[19]。本文针对高产优质滇龙胆准确、高效鉴别的生产需求,以877株滇龙胆有效成分产量性状数据为基础,利用PLS-DA、OPLS-DA、RF 3种算法建立分级鉴别模型。结果发现,RF算法分类正确率最高,模型性能最优。作为目前分类效果较好的集成算法,RF已被广泛应用于植物遗传育种、样品分类、表型多样性分析等[34-36]。与目前药用植物分类建模报道较多的PLS-DA、OPLS-DA等算法相比,RF算法能够较好地克服植物根、茎、叶产量性状数据间存在的共线性问题,有效提高了模型预测精度和稳定性[37]。本研究首次将有效成分产量性状与RF算法相结合,应用于滇龙胆优质高产种源的鉴别,并取得了较好的结果。随着后续建模样品数量及表型数据的增加,模型还可进一步优化完善,最终服务于滇龙胆药材的生产。
1 如需查阅附件内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。2 1 ~ 3)如需查阅附件内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。3 1 ~ 3)如需查阅附件内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。4 如需查阅附件内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。5 1 ~ 3)如需查阅附件内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。6 如需查阅附件内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。 -
![]()
图 2 滇龙胆样品偏最小二乘-判别分析得分图(A)与VIP分析图(B)
Figure 2. Loading plot based on PLS-DA (A) and VIP analysis (B) of Gentiana rigescens samples
![]()
图 3 样品偏最小二乘-判别分析得分图(A)与VIP分析图(B)
Figure 3. Loading plot of PLS-DA (A) and VIP analysis (B)
![]()
图 6 随机森林模型ntree(A)与mtry(B)的参数筛选
Figure 6. ntree (A) and mtry (B) screening of RF model
表 1 滇龙胆有效成分产量性状的多样性指数
Table 1 Shannon diversity index of yield traits of active ingredients in Gentiana rigescens
产量性状
Yield traitShannon多样性指数(I) Shannon diversity index 根Root 茎Stem 叶Leaf 马钱苷酸 1.51 0.94 0.73 6'-O-β-D-葡萄糖基龙胆苦苷 1.39 1.15 1.02 獐牙菜苦苷 1.53 1.20 1.09 龙胆苦苷 1.64 1.35 1.05 当药苷 1.37 1.56 0.73 异荭草素 − 1.08 1.05  下载: 导出CSV
下载: 导出CSV
表 2 基于滇龙胆17个产量性状的主成分分析
Table 2 Principal component analysis results based on 17 yield traits of Gentiana rigescens
成分
Component初始特征值Eigenvalue 特征值
Eigenvalue方差贡献率
Contribution of variance / %累积方差贡献率
Cumulative contribution / %1 7.619 44.816 44.816 2 2.700 15.885 60.700 3 1.602 9.423 70.123 4 1.187 6.982 77.105 5 0.781 4.592 81.697
下载: 导出CSV
表 3 基于PLS-DA算法的分类建模评价结果
Table 3 Evaluation results of classification model based on PLS-DA algorithm
评价指标
Evaluation parameter一级
Class 1 / %二级
Class 2 / %三级
Class 3 / %合计
Total / %训练集正确率 92.00 82.24 89.92 88.05 测试集正确率 92.06 86.11 94.05 90.74 注:一级为优质种源,二级为普通种源,三级为劣质种源。下同。 Notes: Classes 1, 2, and 3 refer to high-quality, moderate-quality, and low-quality germplasms, respectively. Same below.
下载: 导出CSV
表 4 基于OPLS-DA算法的滇龙胆分类模型
Table 4 Evaluation results of classification model based on OPLS-DA algorithm
评价指标
Evaluation parameter一级
Class 1 / %二级
Class 2 / %三级
Class 3 / %合计
Total / %训练集正确率 92.48 82.24 89.44 88.05 测试集正确率 92.86 86.51 93.65 91.01
下载: 导出CSV
表 5 基于随机森林算法建立的滇龙胆分类模型
Table 5 Evaluation results of classification model based on RF algorithm
评价指标
Evaluation parameter一级
Class 1 / %二级
Class 2 / %三级
Class 3 / %合计
Total / %训练集正确率 95.37 91.21 95.85 94.14 测试集正确率 96.43 92.06 95.63 94.71
下载: 导出CSV
-
[1] 徐燕玲,王振宇,杨淑达,陆露. 进化生态学在药用植物种质资源评价中的应用与展望[J]. 中草药,2021,52(5):1221−1233. Xu YL,Wang ZY,Yang SD,Lu L. Application and prospect of evolutionary ecology in evaluation of germplasm resources of medicinal plants[J]. Chinese Traditional and Herbal Drugs,2021,52 (5):1221−1233.
[2] 闫婕,彭成,裴瑾,马云桐,高继海,陈江. 国家中药种质资源库的建设思路与发展策略[J]. 成都中医药大学学报,2021,44(1):14−19,31. Yan J,Peng C,Pei J,Ma YT,Gao JH,Chen J. Construction ideas and development strategies of the state bank of Chinese drug germplasm resources[J]. Journal of Chengdu University of Traditional Chinese Medicine,2021,44 (1):14−19,31.
[3] Wang WL,Xu JF,Fang HY,Li ZJ,Li MH. Advances and challenges in medicinal plant breeding[J]. Plant Sci,2020,298:110573. doi: 10.1016/j.plantsci.2020.110573
[4] 刘义飞,胡志刚,黄必胜,陈士林. 组学技术在中药种质资源遗传评价与创新中的应用[J]. 世界科学技术-中医药现代化,2022,24(4):1315−1324. Liu YF,Hu ZG,Huang BS,Chen SL. Application of omics approaches in genetic evaluation and innovation of traditional Chinese medicine germplasm resources[J]. Modernization of Traditional Chinese Medicine and Materia Medica-World Science and Technology,2022,24 (4):1315−1324.
[5] 孟祥才,于鹏程,何录文,关瑜. 从社会发展角度探讨中药资源的历史、现在与未来[J]. 中草药,2022,53(16):5235−5244. Meng XC,Yu PC,He LW,Guan Y. Discussion on past,present and future of traditional Chinese medicine resources based on social development level[J]. Chinese Traditional and Herbal Drugs,2022,53 (16):5235−5244.
[6] 李智敏,刘莉,李晚谊,张金渝,金航. 滇龙胆的药用资源研究与开发进展[J]. 云南大学学报(自然科学版),2009,31(S1):485−487,491. Li ZM,Liu L,Li WY,Zhang JY,Jin H. Progress on research and development of G. rigescens as a raw material[J]. Journal of Yunnan University,2009,31 (S1):485−487,491.
[7] 国家药典委员会. 中华人民共和国药典: 一部[M]. 北京: 中国医药科技出版社, 2020: 99-100. [8] 褚博文,张霁,李智敏,王元忠. 滇龙胆化学成分和药理作用研究进展[J]. 中国实验方剂学杂志,2016,22(13):213−222. Chu BW,Zhang J,Li ZM,Wang YZ. Research advances in chemical constituents and pharmacological activity from Gentiana rigescens[J]. Chinese Journal of Experimental Traditional Medical Formulae,2016,22 (13):213−222.
[9] 张琳,罗智渊,冯丽丽,李晓东,李海峰. 滇龙胆花药材质量标准及品质评价的研究[J]. 中国药学杂志,2014,49(16):1451−1457. Zhang L,Luo ZY,Feng LL,Li XD,Li HF. Quality evaluation of flowers of Gentiana rigescens Franch.[J]. Chinese Pharmaceutical Journal,2014,49 (16):1451−1457.
[10] 吕伟奇,张霁,左智天,王元忠,张庆芝. 基于灰色关联度分析法的滇龙胆质量评价[J]. 中国实验方剂学杂志,2017,23(13):66−73. Lü WQ,Zhang J,Zuo ZT,Wang YZ,Zhang QZ. Quality evaluation of Gentiana rigescens by grey relational analysis method[J]. Chinese Journal of Experimental Traditional Medical Formulae,2017,23 (13):66−73.
[11] 杨天梅,杨美权,杨绍兵,许宗亮,杨维泽,等. 坚龙胆药材产品质量分级标准研究[J]. 中药材,2018,41(3):648−651. [12] Pan Y,Zhao YL,Zhang J,Li WY,Wang YZ. Phytochemistry and pharmacological activities of the genus Gentiana (Gentianaceae)[J]. Chem Biodivers,2016,13 (2):107−150. doi: 10.1002/cbdv.201500333
[13] Shen T,Yu H,Wang YZ. Assessing geographical origin of Gentiana rigescens using untargeted chromatographic fingerprint,data fusion and chemometrics[J]. Molecules,2019,24 (14):2562. doi: 10.3390/molecules24142562
[14] Shen T,Yu H,Wang YZ. Assessing the impacts of climate change and habitat suitability on the distribution and quality of medicinal plant using multiple information integration:take Gentiana rigescens as an example[J]. Ecol Indic,2021,123:107376. doi: 10.1016/j.ecolind.2021.107376
[15] 康传志,张燕,王升,万修福,蒋靖怡,等. 基于多个利益相关方的中药生态农业经济效益分析[J]. 中国中药杂志,2021,46(8):1858−1863. Kang CZ,Zhang Y,Wang S,Wan XF,Jiang JY,et al. Analysis of economic benefits of Chinese medicine eco-agriculture based on multiple stakeholders[J]. China Journal of Chinese Materia Medica,2021,46 (8):1858−1863.
[16] 孟祥才,杜虹韦,魏文峰,霍金海. 中药资源发展存在的问题与对策[J]. 中草药,2018,49(16):3735−3741. Meng XC,Du HW,Wei WF,Huo JH. Problems and countermeasures in development of Chinese materia medica resource[J]. Chinese Traditional and Herbal Drugs,2018,49 (16):3735−3741.
[17] 段金廒,宿树兰,严辉,郭盛,刘睿,等. 2016−2020年我国中药资源学学科建设及科学研究进展与展望[J]. 中草药,2021,52(17):5151−5165. Duan JA,Su SL,Yan H,Guo S,Liu R,et al. Discipline development and research progress of Chinese medicinal material resources from 2016 to 2020[J]. Chinese Traditional and Herbal Drugs,2021,52 (17):5151−5165.
[18] 贾瑞玲,赵小琴,南铭,陈富,刘彦明,等. 64份苦荞种质资源农艺性状遗传多样性分析与综合评价[J]. 作物杂志,2021(3):19−27. Jia RL,Zhao XQ,Nan M,Chen F,Liu YM,et al. Genetic diversity analysis and comprehensive assessment of agronomic traits of 64 tartary buckwheat germplasms[J]. Crops,2021 (3):19−27.
[19] Coelho C,Figueredo G,Lafarge C,Bou-Maroun E,Flahaut S. Mid-infrared spectroscopy combined with multivariate analysis and machine-learning:a powerful tool to simultaneously assess geographical origin,growing conditions and bitter content in Gentiana lutea roots[J]. Ind Crops Prod,2022,187:115349. doi: 10.1016/j.indcrop.2022.115349
[20] 宋雪彬,高康,黄河,刘芷兰,戴思兰,嵇彧. 中国传统大菊叶片形态的数量化定义与分类[J]. 植物学报,2021,56(1):10−24. Song XB,Gao K,Huang H,Liu ZL,Dai SL,Ji Y. Quantitative definition and classification of leaves in large-flowered Chinese chrysanthemum based on the morphological traits[J]. Chinese Bulletin of Botany,2021,56 (1):10−24.
[21] Liu CY,Yang ZY,Hu YG. Drought resistance of wheat alien chromosome addition lines evaluated by membership function value based on multiple traits and drought resistance index of grain yield[J]. Field Crops Res,2015,179:103−112. doi: 10.1016/j.fcr.2015.04.016
[22] 李红宇,李逸,司洋,杜春颖,周雪松,等. 北方粳稻耐盐碱相关性状主成分分析及综合评价[J]. 核农学报,2020,34(8):1862−1871. Li HY,Li Y,Si Yang,Du CY,Zhou XS,et al. Principal component analysis and comprehensive evaluation of saline-alkaline tolerance related traits of northern Japonica rice[J]. Journal of Nuclear Agricultural Sciences,2020,34 (8):1862−1871.
[23] 王元忠,沈涛,张金渝. 滇重楼及其近缘种的表型变异与资源评价[J]. 热带作物学报,2021,42(9):2535−2541. Wang YZ,Shen T,Zhang JY. Phenotypic variation and resource evaluation of Paris polyphylla var. yunnanensis and its relative species[J]. Chinese Journal of Tropical Crops,2021,42 (9):2535−2541.
[24] Wang CY,Tang L,Li L,Zhou Q,Li YJ,et al. Geographic authentication of Eucommia ulmoides leaves using multivariate analysis and preliminary study on the compositional response to environment[J]. Front Plant Sci,2020,11:79. doi: 10.3389/fpls.2020.00079
[25] Chen XJ,Min DH,Yasir TA,Hu YG. Evaluation of 14 morphological,yield-related and physiological traits as indicators of drought tolerance in Chinese winter bread wheat revealed by analysis of the membership function value of drought tolerance (MFVD)[J]. Field Crops Res,2012,137:195−201. doi: 10.1016/j.fcr.2012.09.008
[26] 刘思辰,曹晓宁,温琪汾,王海岗,田翔,等. 山西谷子地方品种农艺性状和品质性状的综合评价[J]. 中国农业科学,2020,53(11):2137−2148. Liu SC,Cao XN,Wen QF,Wang HG,Tian X,et al. Comprehensive evaluation of agronomic traits and quality traits of foxtail millet landrace in Shanxi[J]. Scientia Agricultura Sinica,2020,53 (11):2137−2148.
[27] 潘晓雪,胡明瑜,王忠伟,吴红,雷开荣. 不同水稻种质资源重要农艺性状与发芽期耐寒性鉴定研究[J]. 作物杂志,2021,37(1):47−53. Pan XX,Hu MY,Wang ZW,Wu H,Lei KR. Evaluation of agronomic traits and cold tolerance at germination stage in rice (Oryza sativa L.) germplasms[J]. Crops,2021,37 (1):47−53.
[28] Aneley GM, Haas M, Köhl K. LIDAR-Based phenotyping for drought response and drought tolerance in potato[J]. Potato Res, 2022, doi: 10.1007/s11540-022-09567-8.
[29] Li L,Li ZM,Wang YZ. A method of two-dimensional correlation spectroscopy combined with residual neural network for comparison and differentiation of medicinal plants raw materials superior to traditional machine learning:a case study on Eucommia ulmoides leaves[J]. Plant Methods,2022,18 (1):102. doi: 10.1186/s13007-022-00935-6
[30] Jiang JL,Johansen K,Stanschewski CS,Wellman G,Mousa MAA,et al. Phenotyping a diversity panel of quinoa using UAV-retrieved leaf area index,SPAD-based chlorophyll and a random forest approach[J]. Precis Agric,2022,23 (3):961−983. doi: 10.1007/s11119-021-09870-3
[31] Wu Z,Zhao YL,Zhang J,Wang YZ. Quality assessment of Gentiana rigescens from different geographical origins using FT-IR spectroscopy combined with HPLC[J]. Molecules,2017,22 (7):1238. doi: 10.3390/molecules22071238
[32] Zhao YL,Yuan TJ,Zhang J,Wang YZ. Geographic origin identification and rapid determination of four constituents of Gentiana rigescens by FTIR combined with chemometrics[J]. J Chemometr,2019,33 (4):e3115. doi: 10.1002/cem.3115
[33] Liu L,Zuo ZT,Xu FR,Wang YZ. Study on quality response to environmental factors and geographical traceability of wild Gentiana rigescens Franch.[J]. Front Plant Sci,2020,11:1128. doi: 10.3389/fpls.2020.01128
[34] Parmley KA,Higgins RH,Ganapathysubramanian B,Sarkar S,Singh AK. Machine learning approach for prescriptive plant breeding[J]. Sci Rep,2019,9 (1):17132. doi: 10.1038/s41598-019-53451-4
[35] Rahaman M,Ahsan M,Chen M. Data-mining techniques for image-based plant phenotypic traits identification and classification[J]. Sci Rep,2019,9:19526. doi: 10.1038/s41598-019-55609-6
[36] Johansen K,Morton MJL,Malbeteau Y,Aragon B,Al-Mashharawi S,et al. Predicting biomass and yield in a tomato phenotyping experiment using UAV imagery and random forest[J]. Front Artif Intel,2020,3:28. doi: 10.3389/frai.2020.00028
[37] 邢晓语,杨秀春,徐斌,金云翔,郭剑,等. 基于随机森林算法的草原地上生物量遥感估算方法研究[J]. 地球信息科学学报,2021,23(7):1312−1324. Xing XY,Yang XC,Xu B,Jin YX,Guo J,et al. Remote sensing estimation of grassland aboveground biomass based on Random Forest[J]. Journal of Geo-Information Science,2021,23 (7):1312−1324.
-
期刊类型引用(1)
1. 龙桂根,黄芝云,吴南生,冯胜,冯超,丁菲,金松松,何利人,王勇,陈玲. 南酸枣种质资源果实性状变异和综合评价. 热带亚热带植物学报. 2024(06): 781-790 .  百度学术
百度学术
其他类型引用(3)
-
其他相关附件
-
PDF格式
沈涛附录 点击下载(515KB)
-

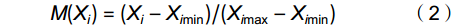

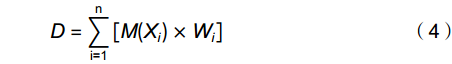
计量
- 文章访问数: 183
- HTML全文浏览量: 34
- PDF下载量: 35
- 被引次数: 4
