
Characterization of leaf phenotypic traits in natural populations of three Polyspora species
-
摘要:
以大头茶属(Polyspora)3种植物的10个天然居群为研究对象,通过多重比较、巢式方差分析、相关性分析、主成分分析和聚类分析等方法,对11个叶表型性状的变异规律进行研究。结果显示:(1)叶表型性状在居群内和居群间均存在极显著差异(P<0.01),平均变异系数为17.03%,不同性状的变异幅度为0.05%~31.37%。叶表型性状在居群内和居群间的方差分量均值分别为180.79和82.63,表明叶表型性状分化程度为居群内高于居群间。(2)叶宽、叶柄长、叶面积、叶基角、叶圆度和叶柄指数为叶表型变异的主导因素;各表型性状之间存在明显的相关性,环境因子对大头茶属植物叶表型性状变异的影响很大;基于叶表型性状可将10个居群分为3类。研究结果可为大头茶属植物的适应性进化和开发利用提供理论依据。
Abstract:Leaf phenotypic variation among 10 populations of three Polyspora species was analyzed using multiple comparisons, nested variance analysis, correlation analysis, principal component analysis (PCA), and cluster analysis. Results revealed: (1) Significant differences in leaf phenotypic traits both within and among Polyspora populations (P<0.01). The average coefficient of variation was 17.03%, with trait-specific variation ranging from 0.05% to 31.37%. Mean variance components for leaf traits were higher within populations (180.79) than among populations (82.63), indicating greater differentiation within populations. (2) Dominant contributors to phenotypic variation included leaf width, petiole length, leaf area, leaf base angle, leaf roundness, and petiole index. Significant correlations among leaf traits were identified, highlighting the strong influence of environmental factors on leaf phenotypic variation. Cluster analysis grouped the 10 populations into three distinct groups based on leaf phenotypic traits. The results provide valuable insights into the adaptive evolution and exploitation of Polyspora species.
-
Keywords:
- Leaf traits /
- Natural population /
- Phenotypic variation /
- Environmental factors /
- Polyspora
-
表型是生物所具有的相对固定的形态和结构特征,表型多样性的形成是基因表达与植物长期适应特定自然环境的综合结果[1-3]。表型性状作为植物遗传多样性的表现形式,其地理变异反映了形态适应性沿着环境梯度的进化模式[4],对于研究物种变异和进化机制具有十分重要的意义。把握物种表型变异的内在规律机制及其对环境因子的响应,可作为探讨植物分类和进化机制的重要参考依据,并为引种驯化和育种改良提供基础[5, 6]。
大头茶属(Polyspora)为山茶科物种丰富度排第2位的类群,主要分布于南亚和东南亚,是印度马来热带雨林-中国亚热带常绿阔叶林的典型代表物种。我国是该属植物的北缘分布区,主要分布于云南、四川、重庆、广西、广东和福建[7-10]。根据杨世雄[7]的分类建议,我国有8种大头茶属植物,分别为大头茶(P. axillaris (Roxburgh ex Ker Gawler) Sweet)、四川大头茶(P. speciosa (Kochs) B. M. Bartholomew & T. L. Ming)、黄药大头茶(P. chrysandra (Cowan) Hu ex B. M. Bartholomew & T. L. Ming)、长果大头茶(P. longicarpa (Hung T. Chang) C. X. Ye ex B. M. Bartholomew & T. L. Ming)、海南大头茶(P. hainanensis (Hung T. Chang) C. X. Ye ex B. M. Bartholomew & T. L. Ming)、天堂大头茶(P. tiantangensis (L. L. Deng & G. S. Fan) S. X. Yang)、越南大头茶(P. tonkinensis (Pit.) T.L. Ming)以及广西大头茶(P. kwangsiensis (H. T. Chang) C. X. Ye)。在《中国生物多样性红色名录-高等植物卷(2020)》[11]和The Red List of Theaceae [12]名录中,其中6种大头茶属植物榜上有名,处于不同程度的濒危状态。
近年来,国内外学者对大头茶属的研究主要集中在分类与分布[7, 8]、遗传分化与基因组结构[13,14]、化学成分[15, 16]、群落结构[17]、地理分布[18]等方面。其表型性状鲜见报道。目前大头茶属的分类还存在分歧或疑问,诸多研究均认为应将越南大头茶、广西大头茶和四川大头茶合并为同一物种。主要依据有:(1)610个直系同源低拷贝核基因的系统发育分析结果显示,四川大头茶和大头茶聚成一个单系[19];(2)本课题组前期研究证明四川大头茶和大头茶的系统发育结果存在核质冲突,四川大头茶、越南大头茶和广西大头茶的叶绿体及核糖体系统发育树均形成高度支持的单系。(3)Flora of China [20]和“中国生物物种名录:2023年度版”(http://www.sp2000.org.cn/)主张将上述3个物种合并,并已做出分类调整。鉴于此,本研究选取这3个物种的10个天然居群,探讨其叶表型变异特征。通过对11个叶表型性状的研究,旨在明确居群间和居群内的表型变异特征及其对地理环境因子的响应机制,探究物种表型特性和表型多样性的影响因素,以期为大头茶属植物的适应性进化研究、种质资源保护、物种分类和可持续开发利用提供参考。
1. 材料与方法
1.1 样品采集
样品采集时间为2020-2022年,采样区域包括云南、四川、重庆、广西、贵州等省市的10个区县。采样植株最小距离为30 m,以尽量降低样本间的亲缘关系。每株随机选取10个成熟、健康且完整的叶片,在东南西北4个方向均匀采集。对于居群植株数量小于20或居群内部表型有明显差异的个体,不受上述取样规则的限制。共采集到10个居群250份叶片样本。在野外采样时,用手持式GPS设备获取经纬度和海拔信息,气候因子从世界气候数据库(https://www.worldclim.org)获取。各居群地理生态因子详见表1。
表 1 大头茶属植物天然居群地理生态因子Table 1. Geo-ecological factors of natural populations of Polyspora序号
No.物种名称(株数)
Species (ind.)采集地点(代码)
Location (Code)纬度
Latitude
(LAT)经度
Longitude
(LONG)海拔
Altitude
(ALT) / m年均温
Mean annual temperature
(MAT)/ ℃年降水
Annual precipitation
(AP) / mm年日照
Annual sunshine
(AS) / h1 P. speciosa(22) 云南昭通盐津县(YJ) 28°2′6.51″N 104°10′44.18″E 946 17.0 676.3 966.0 2 P. speciosa(25) 重庆北碚缙云山(BB) 29°49′47.84″N 106°23′14.43″E 821 18.3 506.0 1 012.2 3 P. speciosa(24) 四川泸州叙永县(XY) 28°17′16.71″N 105°29′21.56″E 686 17.9 1 067.9 1 170.3 4 P. speciosa(21) 贵州遵义习水县(XS) 28°24′46.73″ N 106°36′4.98″E 1 370 17.0 1 933.6 1 269.0 5 P. kwangsiensis(20) 广西来宾金秀县(JX) 24°14′52.93″N 110°12′41.32″E 1 031 20.1 2 086.2 1 413.7 6 P. kwangsiensis(24) 广西桂林恭城县(GC) 24°59′53.27″N 110°56′23.30″E 460 19.9 1 558.1 1 549.1 7 P. kwangsiensis(24) 广西贺州平桂区(PG) 24°37′15.73″N 111°32′34.92″E 617 20.2 1 609.7 1 274.8 8 P. kwangsiensis(16) 广西百色那坡县(NP) 23°17′36.50″N 105°47′29.4″E 1 135 16.5 1 603.7 1 288.3 9 P. kwangsiensis(7) 广西桂林永福县(YF) 24°52′39.36″N 110°09′4.39″E 296 13.6 1 420.6 1 183.7 10 P. tonkinensis(22) 云南红河屏边县(PB) 22°54′38.64″ E 103°42′2.45″ N 2 048 18.8 1 545.6 1 937.3 1.2 表型性状测定
采用游标卡尺与软件相结合的方法测量并计算11个叶表型性状,分别为叶柄长(Petiole length,PL)、叶长(Leaf length,LL)、叶宽(Leaf width,LW)、叶周长(Leaf perimeter,LP)、叶面积(Leaf area,LA)、叶基角(Leaf base angle,LBA)、叶尖角(Leaf tip angle,LTA )、叶柄指数(Petiole index,PI)、叶形指数(Leaf shape index,LSI)、叶圆度(Leaf roundness,LR)、椭圆度(Ellipticalness index,EI)。其中,PL、LL、LW直接用游标卡尺测量,将清晰的扫描图像导入到Digimizer 6.0图像处理软件中测量LP、LA、LTA、LBA等指标,PI、LSI、LR、EI通过以下公式计算。
(1) (2) (3) (4) 1.3 数据处理
采用巢式方差分析检测3种植物叶表型性状在居群间和居群内的差异,线性模型为:Yijn=μ+αi+βj(i)+εijn,Yijn为第i个居群的第j个个体的第n个观测值,μ为总体平均值,αi为第i个居群的效应值(固定),βj(i)为第i个居群内第j个个体的效应值(随机),εijn为第ijn个观测值的试验误差[21]。进行Duncan多重比较和差异性检验,分别计算10个居群11个叶表型性状的平均值(Х)、标准差(SD),再据此计算叶表型分化系数(Vst)和变异系数(CV)。计算公式如下:
(5) (6) 式中,δ2t/s为居群间方差分量,δ2s为居群内方差分量[22]。采用Pearson相关性分析法,探讨叶表型性状之间及其与地理生态因子之间的相关性。在方差分析和多重比较前对数据进行预处理,检验其是否符合正态分布及是否具有方差齐性,若不符合相关要求,则对数据进行转换。基于叶表型性状对10个天然居群进行非加权配对算术平均法(UPGMA)聚类分析,并绘制树状聚类图。采用SPSS 25.0与JASP 0.18.1.0软件相结合进行数据分析与处理,图形绘制利用GraphPad Prism 9软件完成。
2. 结果与分析
2.1 大头茶属植物天然居群叶表型性状特征
多重比较结果显示, 11个叶表型性状在大头茶属3种植物居群间存在显著差异(P<0.05,表2)。其中,PL、LL、LW、LP和LA均为YJ均值最大,JX均值最小。LSI以PB均值最大,JX均值最小。PI以JX均值最大,GC均值最小。LTA、LBA、LR和EI均以YJ均值最小,LTA为GC均值最大,LBA、LR和EI均以JX均值最大。
表 2 大头茶属植物10个天然居群叶表型性状多重比较Table 2. Multiple comparisons of leaf phenotypic traits among 10 natural populations of Polyspora叶表型性状
Leaf
phenotypic
trait居群
PopulationYJ BB XY JX GC PG NP PB XS YF PL 22.33±4.06ab 21.30±4.53bc 20.35±4.77bcd 15.37±4.35g 17.86±3.63defg 17.26±3.29efg 19.72±4.61cde 16.78±3.29fg 24.68±5.58a 18.12±2.62def LL 251.28±21.98a 232.78±37.78b 219.20±36.93bc 165.05±19.96e 208.44±21.88cd 198.99±32.13d 222.36±36.36bc 211.72±26.61cd 208.37±49.64bc 199.34±17.73cd LW 69.45±9.20a 67.87±13.71ab 59.47±8.75c 52.96±11.01d 62.85±5.58bc 61.42±6.66c 61.37±9.60c 51.69±7.87d 57.46±13.30cd 59.95±10.88de LSI 3.66±0.41bc 3.47±0.43bcd 3.71±0.48b 3.25±0.78d 3.33±0.36cd 3.25±0.49d 3.66±0.59bc 4.14±0.49a 3.65±0.43ab 3.63±0.22bc PI 0.09±0.02abc 0.09±0.01abc 0.09±0.02abc 0.09±0.02ab 0.09±0.02bc 0.09±0.02bc 0.09±0.02bc 0.08±0.02c 0.12±0.03c 0.09±0.01d LTA 32.30±7.15e 32.52±9.82e 34.51±13.37de 62.96±23.85b 53.08±14.25c 60.33±20.50bc 42.89±14.81d 36.81±11.42de 50.60±15.33c 41.57±13.80d LBA 36.20±6.25e 55.57±7.04abc 47.21±10.15cd 62.43±22.83a 52.04±14.96bcd 55.19±12.30abc 60.66±19.16ab 45.58±12.13d 46.71±7.92d 47.37±10.17de LP 62.88±6.25a 57.14±9.76b 54.11±8.51bc 40.86±4.74e 50.94±4.56cd 48.21±6.90d 53.75±7.16bc 52.28±6.79c 48.64±10.56d 48.66±5.23cd LA 101.85±20.15a 94.13±25.26bc 79.04±24.07c 54.87±14.26e 80.63±13.18c 76.87±16.88cd 85.68±23.81bc 67.64±18.56d 67.99±30.13de 65.98±10.88e LR 0.32±0.03f 0.36±0.04cd 0.34±0.05de 0.41±0.07b 0.39±0.04bc 0.42±0.05b 0.37±0.06c 0.31±0.03f 0.35±0.05ef 0.35±0.03d EI 0.74±0.03e 0.76±0.06de 0.76±0.03de 0.80±0.03b 0.78±0.02bc 0.80±0.03b 0.79±0.03bc 0.78±0.03cd 0.71±0.13d 0.76±0.02de 注:同行不同小写字母表示差异显著(P<0.05)。PL:叶柄长;LL:叶长;LW:叶宽;LP:叶周长;LA:叶面积;LBA:叶基角;LTA:叶尖角;PI:叶柄指数;LSI:叶形指数;LR:叶圆度;EI:椭圆度。居群缩写为样本采集地代码,其含义与表1相同,下同。 Notes: Different lowercase letters in the same line indicate significant difference (P<0.05). PL: Petiole length; LL: Leaf length; LW: Leaf width; LP: Leaf perimeter; LA: Leaf area; LBA: Leaf base angle; LTA: Leaf tip angle; PI: Petiole index; LSI: Leaf shape index; LR: Leaf roundnes; EI: Ellipticalness index. Abbreviations of population indicate sampling locations as in Table 1. Same below. 2.2 大头茶天然居群叶表型性状变异分析
大头茶属3种植物11个叶表型性状的平均变异系数为0.05%~31.37%(表3),变异系数均值为17.03%。其中,LTA(31.37%)、LA(25.34%)、LBA(23.17%)、PL(20.70%)的变异系数均超20%,说明这些性状特征在居群内部变异较大;EI变异系数仅0.05%,变异程度最小。在居群内部,JX(21.33%)、XS(20.85%)的综合性状变异系数较大,表明居群内部性状变异相对较大,而YF(13.05%)综合性状变异系数最小,表明其性状特征较为稳定。
表 3 大头茶属植物天然居群表型性状变异系数Table 3. Coefficients of variation of phenotypic traits of natural populations of Polyspora居群
Population叶表型性状变异系数Coefficients of variation of leaf phenotypic traits / % PL LL LW LSI PI LTA LBA LP LA LR EI 均值Mean YJ 17.80 8.57 12.98 11.11 20.17 21.69 16.93 9.75 19.39 7.75 0.04 13.29 BB 20.84 15.90 19.80 12.25 17.93 29.59 12.41 16.74 26.29 11.07 0.07 16.63 XY 22.95 16.51 14.41 12.67 19.67 37.97 21.06 15.4 29.83 14.43 0.04 18.63 JX 27.72 11.85 20.38 23.58 24.26 37.11 35.84 11.36 25.47 17.01 0.04 21.33 GC 19.92 10.29 8.70 10.50 17.40 26.30 28.17 8.77 16.02 9.92 0.02 14.18 PG 18.67 15.82 10.62 14.80 17.77 33.29 21.87 14.02 21.52 11.66 0.03 16.37 NP 22.89 16.02 15.32 15.79 20.33 33.84 30.94 13.05 27.23 15.88 0.03 19.21 PB 19.22 12.31 14.92 11.67 19.62 30.39 26.07 12.72 26.89 11.03 0.04 16.81 XS 22.60 23.82 23.15 11.80 21.04 30.30 16.96 21.70 44.32 13.53 0.18 20.85 YF 14.45 8.89 8.99 6.04 14.16 33.20 21.48 10.75 16.48 9.06 0.03 13.05 均值 20.70 14.00 14.93 13.02 19.24 31.37 23.17 13.43 25.34 12.13 0.05 17.03 2.3 天然居群叶表型性状的变异来源
巢式方差分析结果显示(表4),大头茶属3种植物11个叶表型性状在居群间存在极显著差异(P<0.01)。居群内和居群间的方差分量分别占71.20%和28.80%,表明居群内的分化程度高于居群间。表型分化系数变化范围为0~38.00%(表5),分化系数较高的性状有LP(38%)、LTA(34.38%)、LR(33.33%)、EI(33.33%)、LL(32.52%)、PL(30.00%)和LA(30.00%),说明这些叶表型性状在居群间分化较大,居群间多样性比居群内丰富。LSI(20.60%)、LBA(23.67%)在居群间分化较小,相对稳定。PI的表型分化系数为0,说明该性状在不同居群间没有差异。11个表型性状的平均分化系数为27.42%,即大头茶植物叶表型变异在居群间和居群内的贡献率分别为27.42%和72.58%,居群内变异是大头茶属植物叶表型性状变异的主要来源。
表 4 大头茶属植物居群表型性状的巢式方差分析Table 4. Nested variance analysis of phenotypic traits of populations of Polyspora表型性状
Phenotypic trait均方Mean square F值 F value 居群间
Inter-population居群内
Intra-population居群间
Inter-population居群内
Intra-populationPL 202.340 8.760 11.111** 0.481 LL 13036.751 698.822 12.626** 0.677 LW 871.434 53.598 9.250** 0.569 LSI 1.780 0.175 7.226** 0.710 PI 0.003 0 9.233** 1.035 LTA 3 234.154 151.352 13.586** 0.636 LBA 1 547.572 90.130 8.298** 0.483 LP 871.083 31.426 15.609** 0.563 LA 4 947.772 268.632 11.261** 0.611 LR 0.034 0.002 15.320** 0.880 EI 0.019 0.002 7.377** 0.850 Note: **, P<0.01. 表 5 大头茶属植物叶表型性状的方差分量与表型分化系数Table 5. Variance components and phenotypic differentiation coefficients of leaf phenotypic traits in Polyspora表型性状
Phenotypic trait方差分量
Variance component方差分量百分比
Percentage of variance components / %表型分化系数
Phenotypic differentiation
coefficient / %居群间
Inter-population居群内
Intra-population随机误差
Random errors居群间
Inter-population居群内
Intra-populationPL 7.403 17.266 4.967 30.01 69.99 30.00 LL 481.503 999.186 38.480 32.52 67.48 32.52 LW 31.252 90.147 11.018 25.74 74.26 25.74 LSI 0.062 0.239 0.549 20.49 79.51 20.60 PI 0 0 0.021 24.70 75.30 0 LTA 120.191 229.373 18.697 34.38 65.62 34.38 LBA 54.829 176.858 15.221 23.66 76.34 23.67 LP 32.709 53.370 9.278 38.00 62.00 38.00 LA 181.019 422.297 24.562 30.00 70.00 30.00 LR 0.001 0.002 0.059 36.72 63.28 33.33 EI 0.001 0.002 0.056 20.61 79.39 33.33 均值 82.630 180.790 11.170 28.80 71.20 27.42 2.4 叶表型性状的主成分分析
采用SPSS 25.0软件对大头茶属3种植物11个叶表型性状进行主成分分析,选取特征值大于1的3个主成分,结果见表6。3个主成分的贡献率分别为39.712%、23.778%、16.738%,累计贡献率达80.228%,其中,第1主成分贡献率最高,LW和LA载荷度最高,均超过0.9,第2主成分中LBA和LR的载荷度最高,第3主成分中PL和PI的载荷度最高。第1主成分主要反映的是叶片大小性状,第2、3主成分反映的分别是叶片形态和叶柄性状。综合分析表明,LW、PL、LA、LBA、LR和PI是大头茶属3种植物叶表型变异的主导因素。
表 6 大头茶属植物叶表型性状的主成分分析Table 6. Principal component analysis of leaf phenotypic traits of Polyspora叶表型性状
Leaf phenotypic trait主成分Principal component PC1 PC2 PC3 PL 0.389 −0.151 0.868 LL 0.873 −0.427 0.083 LW 0.915 0.320 0.096 LSI −0.073 −0.918 −0.019 PI −0.278 0.174 0.897 LTA −0.363 0.673 0.006 LBA −0.053 0.686 0.003 LP 0.897 −0.371 0.042 LA 0.982 −0.002 −0.048 LR −0.018 0.896 −0.210 EI −0.115 0.312 −0.559 特征值 4.368 2.616 1.841 贡献率 / % 39.712 23.778 16.738 累计贡献率 / % 39.712 63.489 80.228 注:粗体表示该性状在PCA轴上的最大载荷。 Note: Bold indicates maximum load on PCA axis for the trait. 2.5 叶表型性状相关性分析
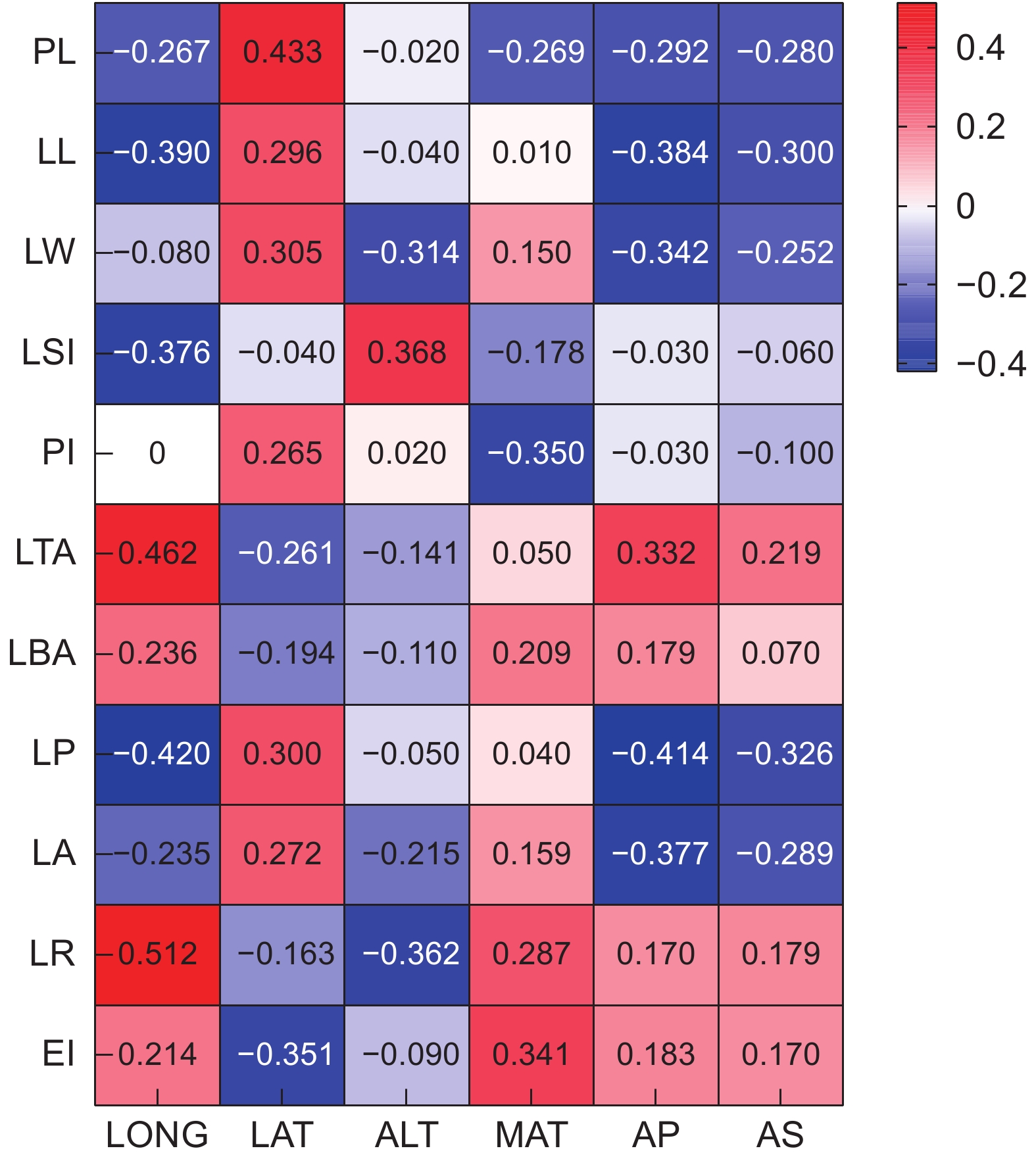
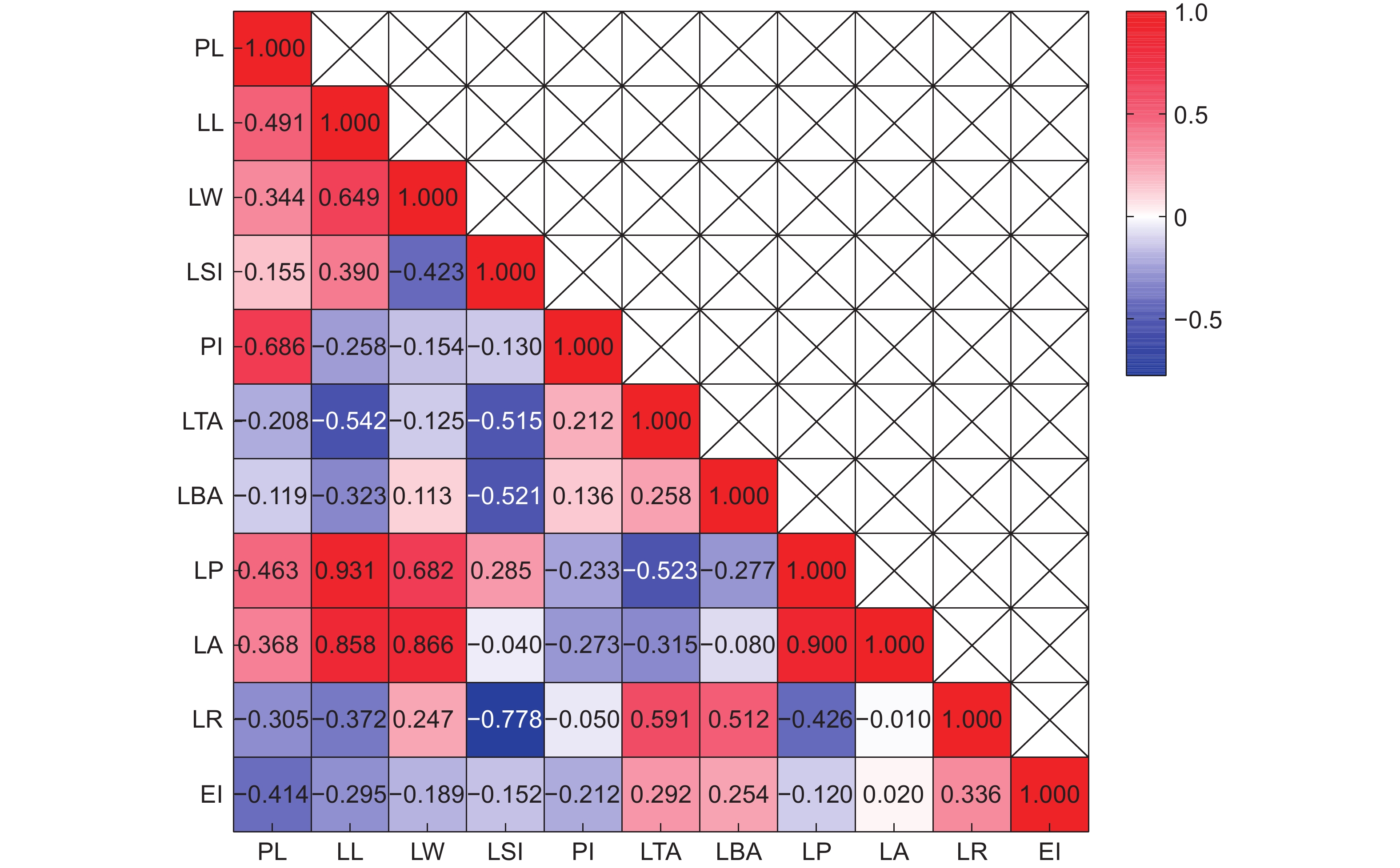
Pearson相关性分析结果表明(图1),PL与除LBA之外的所有性状均呈显著相关(P<0.01)。LL分别与LW、LSI、LP、LA显著正相关(P<0.01);LW分别与LP、LA、LR显著正相关;LSI与LP显著正相关;PI分别与LTA、LBA显著正相关;LTA分别与LBA、LR、EI显著正相关;LBA分别与LR、EI显著正相关。性状之间的相关性表明,LL、LW、LA、LBA是影响叶片表型的关键性状。叶表型性状与地理环境因子的相关性分析结果显示,大部分叶表型性状与地理环境因子间存在显著相关性(图2)。
![]() 图 1 大头茶属植物叶表型性状相关性热图PL:叶柄长;LL:叶长;LW:叶宽;LP:叶周长;LA:叶面积;LBA:叶基角;LTA:叶尖角;PI:叶柄指数;LSI:叶形指数;LR:叶圆度;EI:椭圆度。下同。Figure 1. Correlation heatmap of leaf phenotypic traits in PolysporaPL: Petiole length; LL: Leaf length; LW: Leaf width; LP: Leaf perimeter; LA: Leaf area; LBA: Leaf base angle; LTA: Leaf tip angle; PI: Petiole index; LSI: Leaf shape index; LR: Leaf roundnes; EI: Ellipticalness index. Same below.
图 1 大头茶属植物叶表型性状相关性热图PL:叶柄长;LL:叶长;LW:叶宽;LP:叶周长;LA:叶面积;LBA:叶基角;LTA:叶尖角;PI:叶柄指数;LSI:叶形指数;LR:叶圆度;EI:椭圆度。下同。Figure 1. Correlation heatmap of leaf phenotypic traits in PolysporaPL: Petiole length; LL: Leaf length; LW: Leaf width; LP: Leaf perimeter; LA: Leaf area; LBA: Leaf base angle; LTA: Leaf tip angle; PI: Petiole index; LSI: Leaf shape index; LR: Leaf roundnes; EI: Ellipticalness index. Same below.![]() 图 2 大头茶属植物叶表型性状与环境因子间的相关性分析Figure 2. Pearson correlation analysis of phenotypic traits and environmental factors in Polyspora
图 2 大头茶属植物叶表型性状与环境因子间的相关性分析Figure 2. Pearson correlation analysis of phenotypic traits and environmental factors in Polyspora2.6 大头茶属植物叶表型性状的地理聚类分析
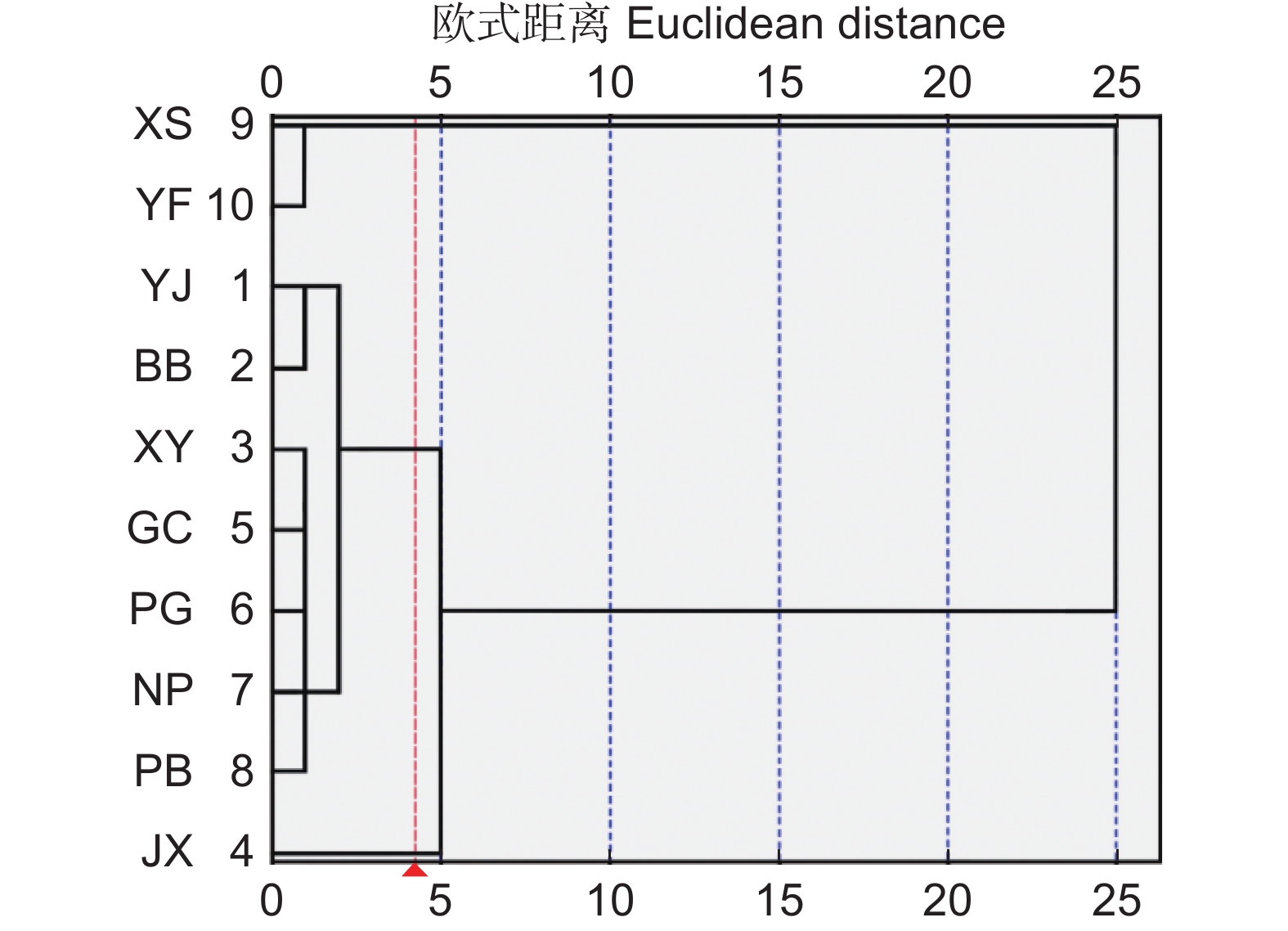
对表型性状特征数据进行标准化处理后,进行UPGMA聚类分析。结果显示,以欧式距离4为阈值,可将10个天然居群分为3类。第1类包括XS和YF;第2类仅包括JX;其余7个居群为第3类(图3)。聚类分析结果表明,大头茶属3种植物叶表型性状特征并没有按照地理距离而聚类,也没有按照物种而聚类,表型性状的变异存在不连续性。
![]() 图 3 基于叶表型性状的UPGMA聚类图Figure 3. UPGMA cluster analysis tree based on leaf phenotypic traits
图 3 基于叶表型性状的UPGMA聚类图Figure 3. UPGMA cluster analysis tree based on leaf phenotypic traits3. 讨论
3.1 大头茶属植物居群叶表型的变异来源
表型性状是遗传多样性和环境多样性综合作用的结果和直观体现。对叶表型性状进行居群差异及地理变异规律进行研究,可在一定程度上反映物种的遗传变异、分布规律及其对生境的适应机制[23]。本研究发现,大头茶属3种植物的叶表型性状在居群间存在极显著差异(P<0.01),而在居群内的差异不显著,说明遗传多样性和环境异质性对叶表型性状变异均有较大影响,且环境异质性的影响更大。大头茶属3种植物居群内的叶表型变异可能主要源于遗传变异,居群间的叶表型变异则可能主要受环境异质性的影响。这与在河北木蓝(Indigofera bungeana Walp.)[5]中的研究结果一致。而在尾叶樱桃(Cerasus dielsiana (Schneid.) Yü et Li)[24]、珙桐(Davidia involucrata Baill.)[25]、小叶锦鸡儿(Caragana microphylla Lam.)[26]等植物中的研究则发现,叶表型性状在居群间和居群内都存在显著或极显著差异。表型变异的差异说明不同物种的叶表型性状受自身遗传因素和环境因子作用的程度不同。
物种天然居群间的变异能更加真实地反映地理与生殖隔离上的差异。本研究发现,大头茶属3种植物11个叶表型性状的平均分化系数为27.42%,低于尾叶樱桃(30.78%)[24]、珙桐(37.0%)[25]以及小叶锦鸡儿(43.62%)[26],高于红椿(Toona ciliata Roem.)(12.13%)[27]。与上述物种相比,由于分布碎片化和地理隔离等因素的综合影响,大头茶属3种植物居群间的叶表型变异处于中等水平。其中,LTA在各性状中的分化系数(34.38%)最高,其在居群内和居群间的方差分量百分比分别为65.62%和34.38%,表明其变异在居群内占据优势;而PI、LR和EI的表型分化系数最低,表明这些性状较为稳定。叶表型性状在居群内的平均方差分量(71.20%)高于居群间(28.80%),表明居群内是变异的主要来源。本研究结果表明,大头茶属3种植物在不同天然居群间的叶形态变异较小。居群内的叶表型变异大于居群间,其原因可能有三。首先,居群地理范围跨度较大,且涵盖不同的气候类型,生境异质性较为突出,形成了不同的生存压力,使得同一居群内的不同个体发育差异明显,促进了大头茶属植物在居群内产生变异。其次,大头茶的繁育机制、种子传播方式和地理分布等方面的差异,可能也导致了其遗传结构的差异。最后,前期研究认为,这3种植物在分类上应合并为同一物种,因此,从这一角度看,其居群间的叶表型性状平均分化系数和变异特征就有可能处于较低水平。
3.2 大头茶属植物居群叶表型变异特征
变异系数越大说明表型变异程度越丰富,这对于植物提高对不同生境的适应能力十分重要。本研究发现,大头茶属3种植物10个天然居群的变异系数均值为17.03%,变异幅度为0.05%~31.37%,变异系数均值与香椿(Toona sinensis (A. Juss.) Roem.)(20.35%)[21]、珙桐(16.22%)[25]接近,说明大头茶属居群存在较大的遗传变异。PL、LTA、LBA、LA等单个性状的变异程度大于LSI、PI、LR、EI等形状指数,说明形状指数相对稳定。叶表型变异系数最大的为LTA(CV=31.37%),表明该指标的可塑性较强,容易受个体发育水平和环境因子的双重影响。变异系数最小的为EI(CV=0.05%),方差分析在居群间达到极显著差异,而在居群内的差异不显著,说明其在遗传上兼具居群间变异性与物种间稳定性的双重特征,该性状不易随环境的变化而改变。叶表型性状与环境因子的相关性分析及主成分分析均表明,EI对大头茶属植物叶表型性状变异的贡献较小。此外,主成分分析结果显示,LW和LA、LBA和LR、PL和PI分别对PC1、PC2、PC3有着较高的贡献率,说明这些性状在叶表型变异中起着主导作用,在今后的研究中应予以重点关注。
3.3 大头茶属植物居群叶表型的地理变异规律
植物叶片对环境的变化较为敏感,有较强的可塑性,叶表型变异有利于避免逆境带来的伤害,增加对环境的生态适应性。相关性分析结果表明,大头茶属植物部分表型性状间具有较高的相关性,如LL与LP的相关系数达0.931,LW与LA的相关系数达0.866,LL与LA的相关系数为0.858,高于紫斑牡丹(Paeonia rockii (S. G. Haw & Lauener) T. Hong & J. J. Li)(0.819)[28]和麻栎(Quercus acutissima Carr.)(0.747)[29],说明大头茶属的叶表型性状对环境因子的变化更为敏感,各表型性状之间具备较强的相互调节能力。
植物的叶表型性状与LONG/LAT存在相关性,主要通过降水、气温和土壤特性等方面表现出来。LAT的改变主要产生气温差异,LONG的变化主要产生降水差异[6]。但地理环境因素之间的关系十分复杂,通常产生综合作用。相关性分析发现,LL、LSI、LP均与LONG显著负相关,LL、LP、LA均与AP显著负相关,而LTA、PL、LSI、LR与LONG/LAT及ALT正相关。一般而言,随着LAT与ALT的增加,气温逐渐降低,空气相对湿度随之减少,植物的叶片大小和叶柄长度会逐渐减小,叶片厚度和宽度也会逐渐下降,以降低水分的散失,提高温度和水分利用效率,从而适应复杂多变的生境。但本研究结果却显示,大头茶属植物叶片随着海拔和经纬度的升高,叶片和叶柄长度都增加,叶面积变大,与在珙桐[25]、皂荚(Gleditsia sinensis Lam.)[30]等植物中的研究结果不一致。其主要原因可能是不同植物的叶表型性状对环境因子的敏感性不同,除了海拔和经纬度因素外,其他地理环境因素如土壤、气温、降水、坡度、坡向等对大头茶属植物的影响更为明显,但具体原因还有待进一步证实。
聚类分析结果表明,欧式距离为4时可将大头茶属的10个天然居群分为3类,各居群并没有按照地理距离而聚类,与尾叶樱桃[24]、珙桐[25]、小叶锦鸡儿[26]等物种中的研究结果相一致。这表明大头茶植物居群的叶表型性状变异具有不连续性,可能是由于大多生长在地势较为复杂的地区,不同居群受地理空间的隔离,限制了其种子和花粉的近距离传播,导致居群间难以进行基因交流,增加了独立分化的机会。总体而言,大头茶属植物在不同地理分布区受局部复杂多样的生境影响,可通过形态和生理等方面的协调互作,以适应异质性生态环境,这可能是其表型多样性较为丰富的重要原因。
4. 结论
本研究表明,不同居群大头茶属植物发生了明显的表型变异,居群内的叶表型性状分化程度高于居群间。
为进一步阐明大头茶属植物的适应性进化及其与环境的作用机制,建议今后从以下3方面开展研究和探索:(1)由于大头茶属的居群数量普遍较少,后期可采取建立自然保护小区、人工育苗、辅助种子入土萌发等措施,扩大天然居群规模,增强其自我更新能力,以增加基因的杂合度,并提高居群的遗传多样性。(2)今后应观测更为全面的表型数据,加强对居群内和居群间变异的研究,探讨叶、花、果、根等表型变异特征与地理环境因子的关系,将形态性状与功能性状联系起来,重点开展叶片生理性状、结构性状、化学性状等方面的研究,并分析其叶经济谱特性,探讨叶片功能性状与环境的关系。(3)地理环境因素对叶表型的影响不仅反映在形态特征层面,生理生化和分子水平也是表型变异的体现。今后可进一步从生理生化和转录组层面对大头茶属植物的表型性状开展研究,以全面揭示其表型变异特征,为解决该属植物分类中存在的分歧提供更为全面的参考依据。
-
![]()
图 1 大头茶属植物叶表型性状相关性热图
PL:叶柄长;LL:叶长;LW:叶宽;LP:叶周长;LA:叶面积;LBA:叶基角;LTA:叶尖角;PI:叶柄指数;LSI:叶形指数;LR:叶圆度;EI:椭圆度。下同。
Figure 1. Correlation heatmap of leaf phenotypic traits in Polyspora
PL: Petiole length; LL: Leaf length; LW: Leaf width; LP: Leaf perimeter; LA: Leaf area; LBA: Leaf base angle; LTA: Leaf tip angle; PI: Petiole index; LSI: Leaf shape index; LR: Leaf roundnes; EI: Ellipticalness index. Same below.
![]()
图 2 大头茶属植物叶表型性状与环境因子间的相关性分析
Figure 2. Pearson correlation analysis of phenotypic traits and environmental factors in Polyspora
![]()
图 3 基于叶表型性状的UPGMA聚类图
Figure 3. UPGMA cluster analysis tree based on leaf phenotypic traits
表 1 大头茶属植物天然居群地理生态因子
Table 1 Geo-ecological factors of natural populations of Polyspora
序号
No.物种名称(株数)
Species (ind.)采集地点(代码)
Location (Code)纬度
Latitude
(LAT)经度
Longitude
(LONG)海拔
Altitude
(ALT) / m年均温
Mean annual temperature
(MAT)/ ℃年降水
Annual precipitation
(AP) / mm年日照
Annual sunshine
(AS) / h1 P. speciosa(22) 云南昭通盐津县(YJ) 28°2′6.51″N 104°10′44.18″E 946 17.0 676.3 966.0 2 P. speciosa(25) 重庆北碚缙云山(BB) 29°49′47.84″N 106°23′14.43″E 821 18.3 506.0 1 012.2 3 P. speciosa(24) 四川泸州叙永县(XY) 28°17′16.71″N 105°29′21.56″E 686 17.9 1 067.9 1 170.3 4 P. speciosa(21) 贵州遵义习水县(XS) 28°24′46.73″ N 106°36′4.98″E 1 370 17.0 1 933.6 1 269.0 5 P. kwangsiensis(20) 广西来宾金秀县(JX) 24°14′52.93″N 110°12′41.32″E 1 031 20.1 2 086.2 1 413.7 6 P. kwangsiensis(24) 广西桂林恭城县(GC) 24°59′53.27″N 110°56′23.30″E 460 19.9 1 558.1 1 549.1 7 P. kwangsiensis(24) 广西贺州平桂区(PG) 24°37′15.73″N 111°32′34.92″E 617 20.2 1 609.7 1 274.8 8 P. kwangsiensis(16) 广西百色那坡县(NP) 23°17′36.50″N 105°47′29.4″E 1 135 16.5 1 603.7 1 288.3 9 P. kwangsiensis(7) 广西桂林永福县(YF) 24°52′39.36″N 110°09′4.39″E 296 13.6 1 420.6 1 183.7 10 P. tonkinensis(22) 云南红河屏边县(PB) 22°54′38.64″ E 103°42′2.45″ N 2 048 18.8 1 545.6 1 937.3  下载: 导出CSV
下载: 导出CSV
表 2 大头茶属植物10个天然居群叶表型性状多重比较
Table 2 Multiple comparisons of leaf phenotypic traits among 10 natural populations of Polyspora
叶表型性状
Leaf
phenotypic
trait居群
PopulationYJ BB XY JX GC PG NP PB XS YF PL 22.33±4.06ab 21.30±4.53bc 20.35±4.77bcd 15.37±4.35g 17.86±3.63defg 17.26±3.29efg 19.72±4.61cde 16.78±3.29fg 24.68±5.58a 18.12±2.62def LL 251.28±21.98a 232.78±37.78b 219.20±36.93bc 165.05±19.96e 208.44±21.88cd 198.99±32.13d 222.36±36.36bc 211.72±26.61cd 208.37±49.64bc 199.34±17.73cd LW 69.45±9.20a 67.87±13.71ab 59.47±8.75c 52.96±11.01d 62.85±5.58bc 61.42±6.66c 61.37±9.60c 51.69±7.87d 57.46±13.30cd 59.95±10.88de LSI 3.66±0.41bc 3.47±0.43bcd 3.71±0.48b 3.25±0.78d 3.33±0.36cd 3.25±0.49d 3.66±0.59bc 4.14±0.49a 3.65±0.43ab 3.63±0.22bc PI 0.09±0.02abc 0.09±0.01abc 0.09±0.02abc 0.09±0.02ab 0.09±0.02bc 0.09±0.02bc 0.09±0.02bc 0.08±0.02c 0.12±0.03c 0.09±0.01d LTA 32.30±7.15e 32.52±9.82e 34.51±13.37de 62.96±23.85b 53.08±14.25c 60.33±20.50bc 42.89±14.81d 36.81±11.42de 50.60±15.33c 41.57±13.80d LBA 36.20±6.25e 55.57±7.04abc 47.21±10.15cd 62.43±22.83a 52.04±14.96bcd 55.19±12.30abc 60.66±19.16ab 45.58±12.13d 46.71±7.92d 47.37±10.17de LP 62.88±6.25a 57.14±9.76b 54.11±8.51bc 40.86±4.74e 50.94±4.56cd 48.21±6.90d 53.75±7.16bc 52.28±6.79c 48.64±10.56d 48.66±5.23cd LA 101.85±20.15a 94.13±25.26bc 79.04±24.07c 54.87±14.26e 80.63±13.18c 76.87±16.88cd 85.68±23.81bc 67.64±18.56d 67.99±30.13de 65.98±10.88e LR 0.32±0.03f 0.36±0.04cd 0.34±0.05de 0.41±0.07b 0.39±0.04bc 0.42±0.05b 0.37±0.06c 0.31±0.03f 0.35±0.05ef 0.35±0.03d EI 0.74±0.03e 0.76±0.06de 0.76±0.03de 0.80±0.03b 0.78±0.02bc 0.80±0.03b 0.79±0.03bc 0.78±0.03cd 0.71±0.13d 0.76±0.02de 注:同行不同小写字母表示差异显著(P<0.05)。PL:叶柄长;LL:叶长;LW:叶宽;LP:叶周长;LA:叶面积;LBA:叶基角;LTA:叶尖角;PI:叶柄指数;LSI:叶形指数;LR:叶圆度;EI:椭圆度。居群缩写为样本采集地代码,其含义与表1相同,下同。 Notes: Different lowercase letters in the same line indicate significant difference (P<0.05). PL: Petiole length; LL: Leaf length; LW: Leaf width; LP: Leaf perimeter; LA: Leaf area; LBA: Leaf base angle; LTA: Leaf tip angle; PI: Petiole index; LSI: Leaf shape index; LR: Leaf roundnes; EI: Ellipticalness index. Abbreviations of population indicate sampling locations as in Table 1. Same below.
下载: 导出CSV
表 3 大头茶属植物天然居群表型性状变异系数
Table 3 Coefficients of variation of phenotypic traits of natural populations of Polyspora
居群
Population叶表型性状变异系数Coefficients of variation of leaf phenotypic traits / % PL LL LW LSI PI LTA LBA LP LA LR EI 均值Mean YJ 17.80 8.57 12.98 11.11 20.17 21.69 16.93 9.75 19.39 7.75 0.04 13.29 BB 20.84 15.90 19.80 12.25 17.93 29.59 12.41 16.74 26.29 11.07 0.07 16.63 XY 22.95 16.51 14.41 12.67 19.67 37.97 21.06 15.4 29.83 14.43 0.04 18.63 JX 27.72 11.85 20.38 23.58 24.26 37.11 35.84 11.36 25.47 17.01 0.04 21.33 GC 19.92 10.29 8.70 10.50 17.40 26.30 28.17 8.77 16.02 9.92 0.02 14.18 PG 18.67 15.82 10.62 14.80 17.77 33.29 21.87 14.02 21.52 11.66 0.03 16.37 NP 22.89 16.02 15.32 15.79 20.33 33.84 30.94 13.05 27.23 15.88 0.03 19.21 PB 19.22 12.31 14.92 11.67 19.62 30.39 26.07 12.72 26.89 11.03 0.04 16.81 XS 22.60 23.82 23.15 11.80 21.04 30.30 16.96 21.70 44.32 13.53 0.18 20.85 YF 14.45 8.89 8.99 6.04 14.16 33.20 21.48 10.75 16.48 9.06 0.03 13.05 均值 20.70 14.00 14.93 13.02 19.24 31.37 23.17 13.43 25.34 12.13 0.05 17.03
下载: 导出CSV
表 4 大头茶属植物居群表型性状的巢式方差分析
Table 4 Nested variance analysis of phenotypic traits of populations of Polyspora
表型性状
Phenotypic trait均方Mean square F值 F value 居群间
Inter-population居群内
Intra-population居群间
Inter-population居群内
Intra-populationPL 202.340 8.760 11.111** 0.481 LL 13036.751 698.822 12.626** 0.677 LW 871.434 53.598 9.250** 0.569 LSI 1.780 0.175 7.226** 0.710 PI 0.003 0 9.233** 1.035 LTA 3 234.154 151.352 13.586** 0.636 LBA 1 547.572 90.130 8.298** 0.483 LP 871.083 31.426 15.609** 0.563 LA 4 947.772 268.632 11.261** 0.611 LR 0.034 0.002 15.320** 0.880 EI 0.019 0.002 7.377** 0.850 Note: **, P<0.01.
下载: 导出CSV
表 5 大头茶属植物叶表型性状的方差分量与表型分化系数
Table 5 Variance components and phenotypic differentiation coefficients of leaf phenotypic traits in Polyspora
表型性状
Phenotypic trait方差分量
Variance component方差分量百分比
Percentage of variance components / %表型分化系数
Phenotypic differentiation
coefficient / %居群间
Inter-population居群内
Intra-population随机误差
Random errors居群间
Inter-population居群内
Intra-populationPL 7.403 17.266 4.967 30.01 69.99 30.00 LL 481.503 999.186 38.480 32.52 67.48 32.52 LW 31.252 90.147 11.018 25.74 74.26 25.74 LSI 0.062 0.239 0.549 20.49 79.51 20.60 PI 0 0 0.021 24.70 75.30 0 LTA 120.191 229.373 18.697 34.38 65.62 34.38 LBA 54.829 176.858 15.221 23.66 76.34 23.67 LP 32.709 53.370 9.278 38.00 62.00 38.00 LA 181.019 422.297 24.562 30.00 70.00 30.00 LR 0.001 0.002 0.059 36.72 63.28 33.33 EI 0.001 0.002 0.056 20.61 79.39 33.33 均值 82.630 180.790 11.170 28.80 71.20 27.42
下载: 导出CSV
表 6 大头茶属植物叶表型性状的主成分分析
Table 6 Principal component analysis of leaf phenotypic traits of Polyspora
叶表型性状
Leaf phenotypic trait主成分Principal component PC1 PC2 PC3 PL 0.389 −0.151 0.868 LL 0.873 −0.427 0.083 LW 0.915 0.320 0.096 LSI −0.073 −0.918 −0.019 PI −0.278 0.174 0.897 LTA −0.363 0.673 0.006 LBA −0.053 0.686 0.003 LP 0.897 −0.371 0.042 LA 0.982 −0.002 −0.048 LR −0.018 0.896 −0.210 EI −0.115 0.312 −0.559 特征值 4.368 2.616 1.841 贡献率 / % 39.712 23.778 16.738 累计贡献率 / % 39.712 63.489 80.228 注:粗体表示该性状在PCA轴上的最大载荷。 Note: Bold indicates maximum load on PCA axis for the trait.
下载: 导出CSV
-
[1] 张睿,国春策,山红艳,孔宏智. 发育重塑与生物多样性[J]. 生物多样性,2014,22(1):66−71. doi: 10.3724/SP.J.1003.2014.13248 Zhang R,Guo CC,Shan HY,Kong HZ. Developmental repatterning and biodiversity[J]. Biodiversity Science,2014,22(1):66−71. doi: 10.3724/SP.J.1003.2014.13248
[2] Li YG,Liu XH,Ma JW,Zhang XM,Xu LA. Phenotypic variation in Phoebe bournei populations preserved in the primary distribution area[J]. J FR,2018,29(1):35−44.
[3] Morton CV. Botany:Principles of Angiosperm Taxonomy. P. H. Davis and V. H. Heywood. Van Nostrand,Princeton,N. J. ,1963. xx + 556 pp. Illus. $15. [J]. Science,1964,144(3618):531.
[4] Wang ML,Zhang JX,Guo ZP,Guan YZ,Qu G, et al. Morphological variation in Cynodon dactylon (L.) Pers. ,and its relationship with the environment along a longitudinal gradient[J]. Hereditas,2020,157(1):4.
[5] 牛雪婧,聂靖,赵雪利,高信芬. 河北木蓝的叶表型可塑性研究[J]. 植物科学学报,2020,38(1):97−104. doi: 10.11913/PSJ.2095-0837.2020.10097 Niu XJ,Nie J,Zhao XL,Gao XF. Leaf-level phenotypic plasticity of Indigofera bungeana Walp[J]. Plant Science Journal,2020,38(1):97−104. doi: 10.11913/PSJ.2095-0837.2020.10097
[6] 郑孙元,朱弘,金思雨,王梦娟,孙杰,等. 桂花表型变化的环境依赖特征[J]. 南京林业大学学报(自然科学版),2019,43(2):38−46. Zheng SY,Zhu H,Jin SY,Wang MJ,Sun J,et al. Environment-dependent phenotypic variation of Osmanthus fragrans[J]. Journal of Nanjing Forestry University (Natural Sciences Edition),2019,43(2):38−46.
[7] 杨世雄. 国产大头茶属的分类处理[J]. 热带亚热带植物学报,2005,13(4):363−365. doi: 10.3969/j.issn.1005-3395.2005.04.017 Yang SX. Taxonomic treatment of Chinese Polyspora Sweet (Theaceae)[J]. Journal of Tropical and Subtropical Botany,2005,13(4):363−365. doi: 10.3969/j.issn.1005-3395.2005.04.017
[8] 马长乐,李靖,白琼,黄晓霞,程小毛. 云南省乡土树种大头茶属植物资源分布与利用探讨[J]. 黑龙江农业科学,2015(5):78−80. Ma CL,Li J,Bai Q,Huang XX,Cheng XM. Distribution and utilization of the indigenous tree species of genus Polyspora in Yunnan province[J]. Heilongjiang Agricultural Sciences,2015(5):78−80.
[9] 陈新艳. 福建省五种植物新记录[J]. 广西植物,2020,40(8):1127−1131. doi: 10.11931/guihaia.gxzw201901022 Chen XY. New records of five species of plants in Fujian province[J]. Guihaia,2020,40(8):1127−1131. doi: 10.11931/guihaia.gxzw201901022
[10] 樊智丰,韩露,马长乐. 大头茶属植物研究进展[J]. 广西植物,2021,41(10):1755−1766. doi: 10.11931/guihaia.gxzw202010042 Fan ZF,Han L,Ma CL. Research advances of Polyspora Sweet (Theaceae)[J]. Guihaia,2021,41(10):1755−1766. doi: 10.11931/guihaia.gxzw202010042
[11] 中华人民共和国生态环境部,中国科学院. 中国生物多样性红色名录——高等植物卷(2020)[M]. 北京:生态环境部,中国国科学院,2023:795,2356−2357. [12] Beech E,Barstow M,Rivers M. The Red List of Theaceae[M]. Richmond,UK:Botanic Gardens Conservation International,2017:32−37.
[13] Fan ZF,Qian SJ,Zhang YH,Ma CL. Characterization of the complete chloroplast genome of Polyspora tiantangensis (Theaceae),an endemic and endangered species in southwestern China[J]. Mitochondrial DNA Part B,2021,6(3):814−815. doi: 10.1080/23802359.2021.1884013
[14] 高灿,樊智丰,马长乐. 黄药大头茶叶绿体基因组密码子偏好性分析[J]. 西南林业大学学报,2023,43(5):66−76. doi: 10.11929/j.swfu.202203018 Gao C,Fan ZF,Ma CL. Analysis of codon usage bias in chloroplast genome of Polyspora chrysandra[J]. Journal of Southwest Forestry University,2023,43(5):66−76. doi: 10.11929/j.swfu.202203018
[15] Athukoralage PS,Herath HMTB,Deraniyagala SA,Wijesundera RLC,Weerasinghe PA. Antifungal constituent from Gordonia dassanayakei[J]. Fitoterapia,2001,72(5):565−567. doi: 10.1016/S0367-326X(00)00339-7
[16] Fu HZ,Li CJ,Yang JZ,Shen ZF,Zhang DM. Potential anti-inflammatory constituents of the stems of Gordonia chrysandra[J]. J Nat Prod,2011,74(5):1066−1072. doi: 10.1021/np200021f
[17] 穆兵,李茂,杨成华,邓伦秀,潘德权,等. 习水国家级自然保护区四川大头茶群落研究[J]. 种子,2011,30(12):62−66. doi: 10.3969/j.issn.1001-4705.2011.12.018 Mu B,Li M,Yang CH,Deng LX,Pan DQ,et al. Study on Sichuan big head tea community in Xishui national nature reserve[J]. Seed,2011,30(12):62−66. doi: 10.3969/j.issn.1001-4705.2011.12.018
[18] Fan ZF,Zhou BJ,Ma CL,Gao C,Han DN,et al. Impacts of climate change on species distribution patterns of Polyspora Sweet in China[J]. Ecol Evol,2022,12(12):e9516. doi: 10.1002/ece3.9516
[19] Zhang Q,Zhao L,Folk RA,Zhao JL,Zamora NA,et al. Phylotranscriptomics of Theaceae:generic-level relationships,reticulation and whole-genome duplication[J]. Ann Bot,2022,129(4):457−471. doi: 10.1093/aob/mcac007
[20] Ming TL,Bartholomew B. Polyspora,Flora of China 12[M]. Beijing:Science Press,2007:418−419.
[21] 姚程程,王俊臣,胡继文,肖遥,杨桂娟,等. 香椿种质生长及叶部表型性状的遗传变异分析[J]. 植物科学学报,2020,38(1):112−122. doi: 10.11913/PSJ.2095-0837.2020.10112 Yao CC,Wang JC,Hu JW,Xiao Y,Yang GJ,et al. Genetic variation of growth and leaf phenotypic traits of Toona sinensis (A. Juss.) Roem. germplasms[J]. Plant Science Journal,2020,38(1):112−122. doi: 10.11913/PSJ.2095-0837.2020.10112
[22] 彭丽平,成仿云,钟原,徐兴兴,鲜宏利. 凤丹栽培群体的表型变异研究[J]. 植物科学学报,2018,36(2):170−180. doi: 10.11913/PSJ.2095-0837.2018.20170 Peng LP,Cheng FY,Zhong Y,Xu XX,Xian HL. Phenotypic variation in cultivar populations of Paeonia ostii T. Hong et J. X. Zhang[J]. Plant Science Journal,2018,36(2):170−180. doi: 10.11913/PSJ.2095-0837.2018.20170
[23] Sun CW,Wang JW,Duan J,Zhao GC,Weng XH,Jia LM. Association of fruit and seed traits of Sapindus mukorossi germplasm with environmental factors in southern China[J]. Forests,2017,8(12):491. doi: 10.3390/f8120491
[24] 朱弘,朱淑霞,李涌福,伊贤贵,段一凡,等. 尾叶樱桃天然种群叶表型性状变异研究[J]. 植物生态学报,2018,42(12):1168−1178. doi: 10.17521/cjpe.2018.0196 Zhu H,Zhu SX,Li YF,Yi XG,Duan YF,et al. Leaf phenotypic variation in natural populations of Cerasus dielsiana[J]. Chinese Journal of Plant Ecology,2018,42(12):1168−1178. doi: 10.17521/cjpe.2018.0196
[25] 张腾驹,陈小红,康喜坤,刘静. 四川省珙桐天然种群叶表型多样性[J]. 生态学杂志,2019,38(1):35−43. Zhang TJ,Chen XH,Kang XK,Liu J. Phenotypic diversity of leaf morphologic traits of Davidia involucrata natural populations in Sichuan Province[J]. Chinese Journal of Ecology,2019,38(1):35−43.
[26] 艾喆,徐婷婷,周兆娜,马飞. 小叶锦鸡儿天然居群叶形态性状变异研究[J]. 西北植物学报,2020,40(9):1595−1604. doi: 10.7606/j.issn.1000-4025.2020.09.1595 Ai Z,Xu TT,Zhou ZN,Ma F. Leaf morphological trait variations in natural populations of Caragana microphylla[J]. Acta Botanica Boreali-Occidentalia Sinica,2020,40(9):1595−1604. doi: 10.7606/j.issn.1000-4025.2020.09.1595
[27] 汪洋,陈文学,明安觉,雍军,宋丛文,等. 湖北红椿天然种群小叶表型性状变异研究[J]. 植物资源与环境学报,2019,28(2):96−105. doi: 10.3969/j.issn.1674-7895.2019.02.13 Wang Y,Chen WX,Ming AJ,Yong J,Song CW,et al. Study on variation of leaflet phenotypic traits of natural populations of Toona ciliata in Hubei Province[J]. Journal of Plant Resources and Environment,2019,28(2):96−105. doi: 10.3969/j.issn.1674-7895.2019.02.13
[28] 张晓骁,宋超,张延龙,牛立新,张庆雨,等. 秦岭与子午岭地区紫斑牡丹居群表型多样性研究[J]. 园艺学报,2017,44(1):139−150. Zhang XX,Song C,Zhang YL,Niu LX,Zhang QY,et al. Phenotypic diversity of Paeonia rockii populations in Qinling and Ziwuling Mountain areas[J]. Acta Horticulturae Sinica,2017,44(1):139−150.
[29] 张元燕,虞木奎,方炎明. 麻栎不同种源的表型性状变异分析[J]. 植物资源与环境学报,2014,23(3):36−44. doi: 10.3969/j.issn.1674-7895.2014.03.05 Zhang YY,Yu MK,Fang YM. Analysis on phenotypic trait variation of different provenances of Quercus acutissima[J]. Journal of Plant Resources and Environment,2014,23(3):36−44. doi: 10.3969/j.issn.1674-7895.2014.03.05
[30] 李伟,林富荣,郑勇奇,李斌. 皂荚南方天然群体种实表型多样性[J]. 植物生态学报,2013,37(1):61−69. doi: 10.3724/SP.J.1258.2013.00007 Li W,Lin FR,Zheng YQ,Li B. Phenotypic diversity of pods and seeds in natural populations of Gleditsia sinensis in southern China[J]. Chinese Journal of Plant Ecology,2013,37(1):61−69. doi: 10.3724/SP.J.1258.2013.00007



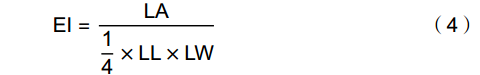


计量
- 文章访问数: 23
- HTML全文浏览量: 1
- PDF下载量: 4
